3. Zonazione della vegetazione marina del Mediterraneo
L'ambiente marino: il Pelago e il Benthos
Prima di descrivere la vegetazione marina sarà bene introdurre, brevemente, le principali suddivisioni in cui è, schematicamente, ripartito l'ambiente marino e gli organismi che si trovano in esso.
Una prima classificazione degli organismi marini va fatta in ragione del fatto che trascorrano la maggior parte della loro esistenza sul fondo marino (benthos), galleggiando nella colonna d'acqua (plancton) o nuotando attivamente (necton).
Questa prima suddivisione corrisponde a quella che viene fatta dell'ambiente marino in dominio pelagico e dominio bentonico. Il primo comprende le acque libere che si estendono dalla superficie alle profondità delle fosse oceaniche. Nel secondo vive l'insieme di organismi che vivono legati al fondale in maniera più o meno diretta.
A seconda della distanza dalla costa il dominio pelagico va suddiviso in provincia neritica, che va dalla linea di costa fino al limite della piattaforma continentale, e in provincia oceanica, che comprende le acque profonde fuori dal limite della piattaforma continentale.
In ragione della profondità e della temperatura la colonna d'acqua è distinta in:
- zona epipelagica o dominio costiero: è la fascia che va dalla superficie fino alla profondità di 50 metri; qui si hanno le maggiori variazioni dei fattori chimico-fisici; questa è anche la zona maggiormente esposta all'inquinamento;
- zona mesopelagica: è la fascia che va da 50 a 200 metri di profondità;
- zona batipelagica: va da 200 a 2500 metri di profondità;
- zona abissopelagica: si estende dai 2500 ai 7000 metri di profondità;
- zona adopelagica: comprende la colonna d'acqua delle fosse abissali.
Un ecosistema comprende tutti le componenti delle comunità di una determinata area geografica e l'ambiente di insediamento: il loro insieme forma il sistema ecologico caratterizzato da complessi circuiti energetici e da molteplici stadi della materia.
Nell'ambiente bentonico si individuano talune comunità con caratteristiche peculiari che costituiscono le biocenosi. Moebius definisce la biocenosi come un raggruppamento di organismi stabilmente insediati in un dato ambiente nel quale vivono e si riproducono raggiungendo una composizione e numero di specie e di individui, tali da non superare le condizioni medie dell'ambiente stesso.
Tutti gli organismi che fanno parte di una biocenosi sono legati tra di loro da rapporti di scambi energetici che ne condizionano la permanenza e sono in equilibrio con le condizioni climatiche ed edafiche del substrato in cui si insediano e che può esser mobile (fanghi, sabbie, ghiaia, ciottoli) o duro (rocce, relitti, manufatti).
L'area geografica che presenta condizioni omogenee che consentono alla biocenosi di svilupparsi è detta biotopo. Quando una limitata area di un biotopo presenta fattori peculiari e parametri adatti per alcuni rappresentanti di comunità differenziate altrove viene detta nicchia ecologica e l'isola di comunità è detta enclave. Se nel biotopo si verifica la predominanza di alcuni fattori che condizionano l'esuberanza di una, o poche, specie senza che questo alteri la composizione qualitativa della biocenosi si ha la formazione della facies. Quando la facies si sviluppa in senso orizzontale si parla di cinture.
Una esuberanza di specie che si ripeta annualmente, nell'ambito della biocenosi, da luogo ad un fenomeno detto periodismo della biocenosi. Gli elementi di una biocenosi si distinguono in caratteristici, compagni e accidentali.
La componente vegetale della biocenosi è limitata nella sua distribuzione in profondità dalla quantità di luce che penetra gli strati d'acqua. In conseguenza di questo fattore nel dominio bentonico si distinguono il sistema fitale ed il sistema afitale. Il primo è quello che si spinge fin dove riescono a vivere i vegetali, il secondo inizia la dove termina il primo e si estende fino alle massime profondità delle fosse abissali.
Nel dominio bentonico si distinguono, inoltre, i piani che comprendono lo spazio verticale caratterizzato da fattori ambientali omogenei e da cenosi di organismi statisticamente e stabilmente correlate.
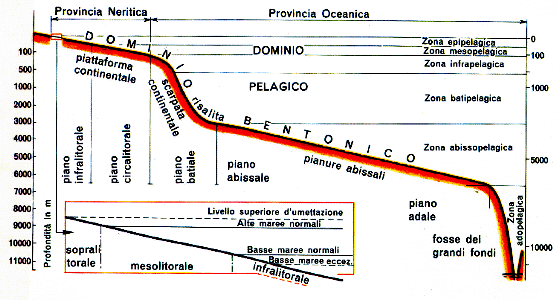
- Nel sistema fitale vi sono cinque piani:
- Adlitorale
- Sopralitorale
- Mesolitorale
- Infralitorale
- Circalitorale
- Il sistema afitale comprende, invece, tre piani:
- Batiale
- Abissale
- Adale
La zonazione del sistema fitale è evidenziata dalla presenza di associazioni vegetali; queste sono definite da una cenosi qualitativamente stabile, ripetitiva in uno spazio sufficiente, determinata da relazioni energetiche, caratterizzata da specie compatibili nell'ecologia, nella corologia e nella sociologia.
Spesso tali associazioni si presentano fisionomicamente come comunità vegetali caratterizzate dalla dominanza di una o poche specie che diventano anche specie guida. I fattori abiotici che determinano il diverso associarsi dei vegetali nei piani sono: l'intensità luminosa, l'intensità e la tipologia dell'idrodinamismo, la tipologia fisica del substrato, la concentrazione in sali nutritivi ed in biostimolanti.
Vegetazione litoranea
La vegetazione litoranea comprende i popolamenti presenti nell'adlitorale, nel sopralitorale e nel mesolitorale. In condizioni di esposizione al moto ondoso frequente ed intenso si evidenzia un'unica zona bionomica: il piano dinamolitorale.
Piano Adlitorale
Questo piano è sotto l'influenza indiretta dell'acqua marina, che arriva sul suolo e sulla vegetazione sotto forma di vento salso o vi si infiltra per capillarità e per fenomeni di penetrazione di origine molto varia. Questo piano, quindi, non è mai raggiunto dall'acqua viva del mare ma solo dalla salsedine.
I litorali possono essere formati da rocce o formazioni sabbiose.Litorale roccioso
La vegetazione delle rupi sul mare è caratterizzata da popolamenti poco densi. Le specie sono poco numerose e coprono scarsamente la superficie delle scogliere tormentate dall'erosione. Gli individui si impiantano nelle fessure, a ridosso di spuntoni di roccia o di grossi massi.
A pochi metri dalla battigia si trovano Ombrellifere e Plumbaginacee. Sulle coste si ha un'associazione tra Chritmum maritimum e varie specie del genere Limonium alle quali si associano alcuni elementi appartenenti alle Composite ed alle Aizacee. Questo tipo di associazione è stata detta Chritmo-Limonietum.
Sui terrazzi rocciosi fino ai 200 metri di altezza sul mare, si impiantano popolamenti più o meno densi di arbusti che costituiscono l'aspetto più degradato dei boschi dell'entroterra. Sulle coste mediterranee dominano le associazioni a Oleo-Lentiscetum e Ceratonietum. Elementi caratteristici della prima associazione sono l'Olivo selvatico ed il Pistacchio selvatico della seconda il Carrubo. Nei versanti più esposti al vento salso si trovano cespugli di Cappero, di Euforbie e rari individui di Quercus ilex (Leccio).
Litorale sabbioso
In particolari situazioni di idrodinamismo o per azione delle barriere formate dalla crescita verticale dei rizomi delle fanerogame marine lungo ampi tratti di litorali marini si formano complessi sabbiosi che per azione del moto ondoso e del vento vengono accumulati a formare dune sabbiose parallele alla linea di costa.
Sulle dune embrionali si impiantano popolamenti effimeri formati da piante pioniere, generalmente a portamento crassulento che consente loro di vivere in ambienti a forte concentrazione salina. Tra questi vegetali pionieri i più comuni sono il Pancratium maritimum, la Cakile maritima. L'Eryngium maritimum, l'Euphorbia paralias e la Crucianella maritima.
L'Ordine fitosociologico in cui sono raggruppate le associazioni psammofile delle regioni mediterranee è detto Ammophiletalia.
Le dune ben stabilizzate sono popolate da Ammophila arenaria e quelle più vicine alla battigia da Agropyrum junceum.
Caratteri comuni ai piani sopralitorale e mesolitorale
I piani sopralitorale e mesolitorale o mediolitorale sono gli unici del sistema fitale a non essere costantemente sommersi. Sono soggetti a forti escursioni diurne e stagionali di temperatura e di salinità. Di conseguenza il fattore umettazione col suo gradiente determina in maniera prioritaria la distribuzione degli organismi in questi due piani.
La vegetazione che si insedia in questo particolarissimo ambiente è caratterizzata dalla presenza funzionale di alghe procariote (Cianoficee). Le Cianoficee infatti svolgono un ruolo determinante nel flusso di energia sia mediante il processo fotosintetico che mediante quello di fissazione dell'azoto. L'importanza delle Cianoficee e di altre alghe endolitiche nel biocarsismo di questi due piani si esplica soprattutto mediante il fenomeno del tranofitismo, che è alla base della circolazione del flusso energetico e dei cicli biogeochimici nel subsistema emerso del sistema fitale.
Anche la capacità diffusa delle Cianoficee di vivere in questi due piani come epi-endofitiche degli altri vegetali contribuisce a rendere disponibile l'azoto da esse fissato sia nel ciclo della materia che nell'efficenza tra questo ciclo ed il flusso dell'energia che lo determina. Le Cianoficee che caratterizzano in maniera preferenziale questi due piani sono: Entophysalis deusta, Calothrix crustacea e Brachytrichia quojii.
Caratteri strutturali ed ecologici fondamentali del sopralitorale
Questo piano vegetazionale coincide con la zona di influenza degli spruzzi causati dall'idrodinamismo superficiale multidirezionale con effetti dirompenti. Corrisponde alla zona olofotica dello schema di Ercegovic e ricade nella zona a idrodinamismo multidirezionale di Riedl.
Dal punto di vista ambientale è l'unico piano costantemente emerso del sistema fitale marino. Presenta forti escursioni termiche, intensa radiazione luminosa, notevoli variazioni della salinità dipendenti dalla evaporazione e dagli apporti di acque meteoriche.
Inferiormente esso è limitato dal livello massimo raggiunto dalle acque per il duplice effetto delle maree e del moto ondoso; superiormente è limitato, sui substrati rocciosi, dal livello raggiunto dagli spruzzi dovuti al moto ondoso e al vento, con una estensione verticale media di sei metri nelle stazioni mediamente battute del Mediterraneo, mentre sui substrati mobili è limitato dal livello raggiunto dalle infiltrazioni di acqua marina nel sedimento.
Sui substrati di roccia calcarea la vegetazione dominante è costituita da Cianoficee e Licheni formanti incrostazioni che si presentano come bande sovrapposte di colore variante dal grigio chiaro al nero. Nella parte più elevata del piano predominano forme epilitiche mentre più in basso, dove maggiore è l'umettazione, ma anche i fenomeni erosivi, predominano forme endolitiche. I substrati di rocce vulcaniche sono molto meno favorevoli all'insediamento delle forme endolitiche e presentano una vegetazione più povera.
Alla luce delle conoscenze attuali, il piano sopralitorale appare popolato da un numero molto ridotto di specie che possono adattarsi alle sue peculiari condizioni ambientali. Con l'eccezione dei licheni Verrucaria maura e V.symbalana, che solo raramente si ritrovano a livelli più bassi, si tratta di Cianoficee che nel sopralitorale sono spesso epilitiche, e che nel mesolitorale, dove sono presenti come componenti abituali di varie associazioni, vivono con più frequenza come endobionti ed endolitiche. Queste specie presentano un accentuato polimorfismo e la loro morfologia è in dipendenza dei fattori ambientali, principalmente l'umidità della roccia, la luce e l'agitazione delle acque.
Aggruppamenti del Sopralitorale
Entophysalidetum deustae Berner 1931
Questa associazione è stata originariamente descritta come Scopulonemetum da Berner (1931) per il Golfo di Marsiglia "in ambienti a luminosità variabile della zona subterrestre, nella fascia soggetta agli spruzzi". Le specie caratteristiche dell'associazione sono: Scopulonema hansgirgianum, Dalmatella buaensis (entrambe sinonimi di Entophysalis deusta), Mastigocoleum testarum, Kyrthutrix dalmatica (= Calothrix crustacea), Verrucaria maura, V.symbalana. L'Autore riporta nella tabella sintetica 13 specie.
Entophysalidetum deustae è, a nostro avviso, una associazione in cui le Cianoficee, in assenza di competizione, diversamente da quanto avviene a livelli inferiori, si diffondono notevolmente e presentano valori di copertura elevati.
Il cambiamento della denominazione è giustificato dal fatto che Scopulonema hansgirgianum è caduto in sinonimia nel 1948 con Entophysalis deusta.
A questo syntaxon possono riferirsi le associazioni proposte da Ercegovic (1932). L'associazione descritta da Ercegovic (1932) come Scopulonemetum hansgirgiani (tab. 11: 215) è da ritenere un sinonimo posteriore. L'autore ha riconosciuto, per i substrati di roccia calcarea del litorale dalmato, una "Zona delle Cianoficee litofite" che si estende verticalmente dal livello delle basse maree sigiziali al livello raggiunto dagli spruzzi di acqua marina per effetto del moto ondoso e dei venti. In essa ha distinto una fascia inferiore, di colore grigiastro o biancastro, in cui predominano le endolite, ed una fascia superiore, di colore bruno-giallastro o nerastro, in cui predominano le epilite. Inoltre, Ercegovic ha riscontrato tale situazione anche in altre coste calcaree del Mediterraneo (da Trieste a Venezia, da Marsiglia a Nizza).
Pozze sopralitorali
Nel sopralitorale si riscontrano degli ambienti alquanto peculiari costituiti da pozze di dimensioni diverse, alimentate sia dagli spruzzi di acqua marina che dalle acque meteoriche. Queste pozze vengono distinte da Feldmann (1937) in "temporanee, a salinità elevata e ricche di sostanze organiche" e in "semipermanenti a salinità ridotta a causa di apporti continui di acque dolci".
Per le pozze temporanee Feldmann (1937) descrive le associazioni a Brachiomonas submarina, a Platymonas tetrathele, a Stephanoptera gracilis, che sono tutte nomina nuda. Cioè le associazioni non sono documentate da rilevamenti. Il variare delle condizioni ecologiche nel corso dell'anno, determina in una stessa pozza il succedersi di popolamenti a composizione specifica molto diversa. Tali popolamenti, per il fatto di essere costantemente immersi, non sono da considerare propriamente sopralitorali. Per le pozze semipermanenti lo stesso Autore descrive una associazione a Rivularia nitida (= Calothrix crustacea) che ha affinità con l'associazione Entophysalidetum deustae.
Caratteri strutturali ed ecologici fondamentali del mesolitorale
Questo piano vegetazionale è compreso tra il sopralitorale, sempre emerso, e l'infralitorale sempre sommerso, ed è caratterizzato da una alternanza più o meno regolare di emersione e di sommersione dipendente principalmente dalla marea e dal moto ondoso. Corrisponde alla zona talantofotica dello schema di Ercegovic e ricade nella zona con idrodinamismo multidirezionale di Riedl (1964, 1971). L'estensione verticale del piano varia da stazione a stazione e dipende dalle escursioni di marea che, in Mediterraneo, hanno valori medi di 20-30 cm e massimi di 1,5-1,8 m. Tale estensione può variare localmente sino a raggiungere oltre 2 metri di ampiezza in rapporto alla conformazione della costa e al moto ondoso.
Sono distinguibili un sottopiano superiore ed uno inferiore, rispettivamente al di sopra e al di sotto del livello medio del mare:
- Il sottopiano superiore presenta un manto vegetale lacunoso e discontinuo con un accentuato periodismo stagionale ed una forte influenza edafica da parte della natura chimica della roccia (silicea o calcarea) e della sua consistenza (compatta o friabile); in esso in condizioni ottimali sono riconoscibili tre orizzonti, evidenziati da tre cinture vegetali.
- Il sottopiano inferiore è caratterizzato da un manto vegetale denso e continuo. Durante la bassa marea, se il mare è calmo, questo sottopiano resta scoperto a lungo, altrimenti viene continuamente umettato per effetto del moto ondoso; in questo sottopiano si distinguono due orizzonti, evidenziati da cinture vegetali, diversificate in differenti situazioni ambientali ed in specifiche regioni biogeografiche.
Aggruppamenti del Mesolitorale
Bangietum atropurpureae Giaccone 1993
L'associazione è presente nell'orizzonte alto del mesolitorale superiore delle stazioni battute su substrati rocciosi. Si sviluppa in ambienti esposti al moto ondoso e raggiunge il pieno sviluppo nei mesi invernali e primaverili. Oltre alla due specie caratteristiche, Bangia atropurpurea e Ulothrix flacca, ha elevati valori di copertura la Cyanophycea tranofita Brachytrichia quojii.
Benchè diversi autori citino associazioni (si tratta di nomina nuda) con le specie caratteristiche sopra menzionate (Feldmann, 1937: associazione a Bangia-Ulothrix; Den Hartog, 1959: Bangieto-Ulothricetum subflaccidae), nessuno di essi pubblica dati quantitativi. In Giaccone (1971), Giaccone et al. (1973) e in Giaccone et al. (1974) sono riportate delle tabelle, ma si tratta di rilievi che ricadono sull'insieme delle cinture mesolitorali comprendenti, oltre al Bangietum atropurpureae, anche altre associazioni.
Porphyretum leucostictae Boudouresque 1971
Specie caratteristiche dell'associazione sono: Porphyra leucosticta, Scytosiphon simplicissimus e Polysiphonia sertularioides. L'autore cita come caratteristica anche Rivularia mesenterica (= Calothrix crustacea), già proposta come caratteristica dell'Entophysalidetum deustae Berner 1931. Inoltre Porphyra umbilicalis e P.linearis, citate dall'Autore nel testo come specie caratteristiche, non sono presenti nei rilevamenti della tabella. Queste due specie sono comunque riscontrabili con frequenza in questa associazione. Questo aggruppamento ha il suo sviluppo ottimale nel periodo invernale e primaverile; è presente nell'orizzonte medio del mesolitorale superiore, in stazioni battute e si colloca tra il Bangietum atropurpureae (in alto) e il Nemalio-Rissoeletum verruculosae (in basso). E' costituita prevalentemente da specie vegetali (ca 90%), tra le quali dominano Neogoniolithon brassica-florida e Scytosiphon simplicissimus; mentre il contingente faunistico è rappresentato soltanto da quattro entità, tra le quali Chthamalus stellatus risulta la specie più importante anche dal punto di vista sociologico. L'analisi della tabella ha evidenziato che il settimo rilevamento è privo di caratteristiche di biocenosi e che in tutti i rilevamenti sono presenti diverse entità (dal 15 % al 40 %) determinate a livello generico, prevalentemente appartenenti al gruppo delle ubiquiste. Il syntaxon si può ritenere valido nonostante la presenza di entità determinate solo a livello generico, in quanto questa carenza è rilevante solo tra il gruppo di specie considerate dall'Autore come ubiquiste e diverse. Ci si augura che in futuro questa associazione sia più compiutamente caratterizzata nel contingente floristico.
Nemalio-Rissoelletum verrucolosae Boudouresque 1971
Le specie caratteristiche di questa associazione sono: Rissoella verruculosa, Nemalion helminthoides e Audouinella nemalionis. L'Autore cita come caratteristica anche Calothrix parasitica (= Calothrix crustacea), già proposta come caratteristica dell'Entophysalidetum deustae Berner 1931. Lo stesso vale per Callithamnion granulatum considerata caratteristica dell'Enteromorphetum compressae Berner 1931. Polysiphonia tenerrima, inoltre, considerata specie caratteristica, non è in tabella. Questo aggruppamento si riscontra durante tutto l'anno ma si sviluppa pienamente nel periodo invernale e primaverile ed è presente nell'orizzonte basso del mesolitorale superiore, in stazioni battute. La tabella ha carattere di omogeneità e le caratteristiche dell'ordine sono più frequenti rispetto a quelle dell'alleanza. L'esame delle correlazioni tra le specie evidenzia un riscontro tra la dominanza di Rissoella verruculosa (ri medio 56%) e il contingente di ordine superiore, mediamente rappresentato a livello di ricoprimento (17%). Inoltre non è privo di significato che in tutti i rilevamenti della tabella si riscontrano diverse determinazioni incomplete (dal 15% al 30%). Sui substrati calcarei Rissoella verruculosa è generalmente assente. Il syntaxon si può ritenere valido con le osservazioni e raccomandazioni riportate per il Porphyretum leucostictae.
Lithophylletum byssoidis Giaccone 1993
Le specie caratteristiche dell'Associazoione sono: Lithophyllum byssoides, Chaetomorpha mediterranea, Laurencia papillosa, Pterocladiella melanoidea, Lophosiphonia cristata, e Taenioma nanum.
L'associazione si inquadra nell'ordine Ralfsietalia verrucosae Giaccone 1993 e nell'alleanza Ralfsion verrucosae Giaccone 1993. Si afferma nel sottopiano inferiore del mesolitorale inferiore del Mediterraneo, con esclusione di alcune aree dell'Egeo, dello Ionio, dell'Adriatico settentrionale e di stazioni di tutto il Mediterraneo con acque dissalate, inquinate o relativamente fredde. In aree e stazioni sfavorevoli può essere vicariato dall'Enteromorphetum compressae Berner 1931. Questa associazione è il tipo nomenclaturale sia dei due syntaxa di ordine superiore appena citati, che della classe Entophysalidetea deustae Giaccone 1993.
Si tratta di una associazione mesofotica, mesoterma, mesoalina e cumatofila di substrato roccioso o duro compatto ed esposto all'azione del moto ondoso e delle escursioni di marea. In condizioni ottimali forma marciapiedi organogeni (trottoir) su substrati suborizzontali e cornici su substrati subverticali. In condizioni meno favorevoli forma cuscini mammellonati sparsi e croste squamate. Il tallo della specie guida è fortemente biocarsificato ad opera di Cyanophyceae tranofite (Brachytrichia quojii, Calothrix crustacea, Entophysalis deusta, E.conferta, Microcoleus lyngbyaceus, Anacystis cyanea) e di Chlorophyceae tranofite (Bulbocoleon pyliferum, Entocladia leptochaete, E.viridis, Phaeophyla dendroides) o comunque da noi rinvenute come prevalentemente endolitiche. Le Cyanophyceae formano un feltro alla base della parte viva del Lithophyllum byssoides e con la fissazione dell'azoto assicurano il ciclo di questo elemento in tutta la formazione organogena.
Lithophylletum byssoidis subass. Lithophylletosum trochanteris Marino, Di Martino & Giaccone 1999.
La specie caratteristica della sub-associazione è: Lithophyllum trochanter.
Giaccone (1968) descrive per molte località dell’Egeo un tipo di vegetazione caratterizzata da Lithophyllum trochanter (Bory) Huvé. Confrontando la bionomia delle associazioni vegetali dell’Egeo e della penisola Maddalena ci sembra che i valori maggiori di temperatura dell’Egeo facciano discendere L.trochanter più tipicamente nella frangia infralitorale e fino a –150 cm di profondità; mentre in Sicilia sud-orientale L.trochanter si sviluppa a partire dall’orizzonte basso del mesolitorale fino a – 80 cm nei substrati sub-verticali. In ogni caso i mammelloni calcarei formati in Sicilia sono di diametro e di altezza inferiori rispetto a quelli dell’Egeo. Secondo la nostra esperienza al di fuori dell’Egeo, L.trochanter forma densi popolamenti solo in alcune stazioni del litorale ibleo. In questo biotopo l’alga calcarea è inserita nell’associazione Lithophylletum byssoidis, come specie differenziale di una sub-associazione che riteniamo nuova per la fitosociologia del Mediterraneo. Nel contingente caratterizzante dell’associazione mancano Taenioma nanum, peraltro presente a Punta Tavernara e Pterocladiella melanoidea, mentre nelle caratteristiche mesolitorali di ordine superiore risultano sempre assenti le Fucophyceae incrostanti, citate in antecedenza. L.trochanter, in questo studio è stato ritrovato a Capo Murro di Porco a –20 cm di profondità associato anche a C.amentacea come in Egeo (Giaccone 1968). Tra le specie accompagnatrici ad alta frequenza troviamo le Cyanophyceae dell’Entophysalidetum deustae, molte specie a ripartizione mesolitorale ed in particolare le specie caratteristiche territoriali della associazione da noi descritta in precedenza come Polysiphonio-Lithophylletum papillosi.
Ceramio-Corallinetum elongatae Pignatti 1962
Le specie caratteristiche dell'associazione sono: Corallina elongata (considerata da noi di ordine superiore), Ceramium ciliatum (senza precisazione della varietà), Ceramium barbatum (= rubrum var. barbatum), Gelidium crinale (= G.pusillum), Antithamnion cruciatum e Grateloupia filicina (con riserva), Ceramium diaphanum var.diaphanum (con riserva). Questa associazione, descritta da Pignatti per la costa veneta come elemento interno di una biocenosi, occupa la porzione più bassa del mesolitorale (denominata in letteratura anche frangia mesolitorale), al limite superiore della frangia infralitorale, ed è emersa solo per brevi periodi. Risulta costituita prevalentemente da vegetali, ma nei rilievi dell'Autore anche la componente faunistica è ben rappresentata (20-23%). Nel litorale veneto l'autore riconosce due subassociazioni, ognuna delle quali comprensiva di due varianti. La subass. ceramietosum, presente nelle stazioni calme, ove si realizza dopo un periodo di tre-quattro anni, è differenziata da Dictyota dichotoma, Enteromorpha linza, Gelidium spathulatum. Invece la subass. corallinetosum, collegata alle stazioni battute, si forma dopo 20-30 anni. In biotopi sciafili corrispondenti si afferma il Phymatolithetum lenormandii Giaccone 1993 Queste due associazioni hanno in comune Corallina elongata che quindi consideriamo anche per questo motivo caratteristica di ordine superiore.
Fucetum virsoidis Pignatti 1962
Le specie caratteristiche dell'associazione sono: Fucus virsoides, Gelidium spathulatum, Phormidium tenue (= Schizothrix calcicola), Enteromorpha flexuosa subsp. flexuosa e Gelidium pulvinatum (= G.pusillum). Le altre specie riportate dall'Autore sono caratteristiche di altre associazioni: Bangia fuscopurpurea (= B.atropurpurea) è caratteristica del Bangietum atropurpureae, Rivularia mesenterica (= Calothrix crustacea) è caratteristica dell'Entophysalidetum deustae. Associazione esclusiva delle coste orientali (dall'Albania alla Slovenia) e settentrionali dell'Adriatico (da Triete a Venezia), essa occupa tutto il piano mesolitorale ed è connessa con le ampie escursioni di marea, con le temperature relativamente basse e con le acque dissalate ed eutrofiche proprie di questo tratto di mare. Si tratta probabilmente di un aggruppamento relitto del bacino della Paratetide. Sulla base della diagnosi originale, risulta costituita prevalentemente da vegetali (86%) mentre la componente faunistica è rappresentata da otto specie (14,%) tra le quali risultano più frequenti: Patella coerulea, Mytilus galloprovincialis, Actinia equina e Balanus sp. La tabella è omogenea e comprende un ricco elenco di caratteristiche. L'Autore rileva anche una facies a Cladophora vagabunda (come penicillata) sviluppata nel mese di settembre. Il successivo studio di Giaccone & Pignatti (1967) sulla vegetazione del golfo di Trieste, ha ulteriormente contribuito a una migliore conoscenza dei popolamenti vegetali a Fucus virsoides, che nel particolare ambiente sono caratterizzati anche dalla presenza di alcune specie rare o assenti nelle coste venete: Catenella caespitosa, Lophosiphonia subadunca, Streblocladia collabens.
Phymatolithetum lenormandii Giaccone 1993
Le specie caratteristiche dell'associazione sono: Phymatolithon lenormandii, Cruoriella armorica (che può discendere anche nei piani sottostanti), Hildenbrandia rubra e Gymnothamnion elegans.
L'associazione si sviluppa in maniera matura sotto il bordo del marciapiede, sulle pareti delle cornici a Lithophyllum byssoides e all'ingresso delle grotte. La tabella dei rilievi riportata mette in evidenza tra le specie caratteristiche anche Cruoriella armorica, Gymnothamnion elegans e Hildenbrandia rubra. Enclaves di questa associazione si trovano in tutto il mesolitorale inferiore, nella frangia infralitorale e nel sottostrato del Cystoseiretum strictae Molinier 1958. Consideriamo affine a questa associazione l'Hildendrandietum prototypi indicato come nomen nudum sia da Giaccone (1965) sia da Boudouresque (1971). Si tratta di situazioni ambientali molto sciafile in cui Phymatolithon lenormandii di fatto scompare.
Enteromorphetum compressae (Berner 1931) Giaccone 1993
Le specie caratteristiche dell'Associazine sono: Enteromorpha compressa, Ralfsia verrucosa (considerata da Molinier (1958) e da noi caratteristica di ordine superiore), Callithamnion granulatum, Cladophora pellucida, Blidingia minima (riportata come Enteromorpha micrococca), Verrucaria maura (che tuttavia appartiene ad un'altra associazione). Oltre a queste caratteristiche considerate dall'autore specie esclusive, ne sono elencate in tabella altre 13.
Questa associazione è stata riscontrata insieme ad altre dell'alleanza Ulvion (Corallinetum e Ceramietum) in una stazione particolare del Golfo di Marsiglia soggetta al moto ondoso ma riparata dall'impatto diretto dei marosi, con apporto di acqua dal largo che attenua lo stato di inquinamento e dissalamento tipiche del golfo. L'Autore considera questa subassociazione, legata come serie laterale al Ceramietum, una formazione aperta e poco ben definita sociologicamente e dipendente all'alleanza Ulvion.
Polysiphonio-Lithiphylletum papillosi Marino, Di Martino & Giaccone 1999
Le specie caratteristiche dell'associazione sono: Lithophyllum papillosum, Polysiphonia opaca, Polysiphonia sertularioides.
Questa associazione era stata già rilevata , ma non descritta a Lampedusa (Scammacca et al., 1993) e si ritrova in tutto il Mediterraneo orientale (Giaccone, 1968). Dal punto di vista bionomico si sviluppa nel mesolitorale superiore, ma si può trovare con specie trasgressive anche nel sottopiano inferiore sui substrati a forte pendenza. Essa vicaria due associazioni che, nel Mediterraneo occidentale, popolano il sottopiano superiore del mesolitorale (Porphyretum leucostictae Boudouresque 1971 e Nemalio-Rissoelletum verruculosae Boudouresque 1971). La specie comune ad entrambi i bacini del Mediterraneo è Polysiphonia sertularioides che si ritrova nel contingente caratterizzante anche del Porphyretum leucostictae. Si tratta evidentemente di una specie che, a giudicare dai valori di ricoprimento e dallo stato riproduttivo, trova condizioni ottimali in questo livello bionomico del Mediterraneo orientale. Mentre P. sertularioides domina nelle porzioni superiori del sottopiano, P. opaca preferisce la parte inferiore. La corallinacea Lithophyllum papillosum si comporta da buona caratteristica territoriale per tutto il piano mesolitorale del Mediterraneo orientale, ma preferisce il sottopiano superiore. Nelle zone di transizione biogeografica si trova associato anche a Lithophyllum lichenoides e ad altre specie a ripartizione mesolitorale caratteristiche di altre associazioni vegetali. Nella tabella hanno elevata frequenza ed alti valori di ricoprimento le Alghe Azzurre, già ritrovate nei rilevamenti illustrati in precedenza per l’Entophysalidetum deustae. Tra le specie occasionali a ripartizione mesolitorale sono presenti in tabella: Callithamnion granulatum, Chondrophycus papillosus, Nemalion helminthoides, Audouinella nemalionis, Ceramium ciliatum v. robustum, Entheromorpha compressa e Taenioma nanum. Mentre è facile inquadrare questa associazione nella classe Entophysalidetea Giaccone 1993 è difficile attribuirla ad ordini ed alleanze già introdotte per il Mediterraneo (Giaccone et al., 1993) Riteniamo che sia necessaria una maggiore conoscenza della vegetazione mesolitorale del Mediterraneo orientale prima di provare ad inquadrare questa associazione in sintaxa mesolitorali di ordine superiore. Peraltro le alghe brune incrostanti Hapalospongidion macrocarpum e Nemoderma tingitanum caratteristiche di sintaxa mesolitorali di ordine superiore sono rare e o assenti nel Mediterraneo orientale (Ribera et al., 1992).
Syntaxa di ordine superiore
Per il piano sopralitorale riconosciamo con Ercegovic i seguenti syntaxa di ordine superiore che aggiorniamo coerentemente nella nomenclatura:
Ord. Entophysalidetalia deustae Ercegovic 1932 nom. mut.
(Den . orig.: Pleurocapsetalia crepidinum Ercegovic 1932: Tab. 9-10: 214).
Car. ord.: Microcoleus lyngbyaceus. Secondo la revisione delle Cyanophyceae, Lyngbya martensiana v. marina è un sinonimo di questa specie.
Tipo nomencleturale: Entophysalidion deustae Ercegovic 1932
All. Entophysalidion deustae Ercegovic 1932 nom. mut.
(Den. orig.: Scopulonemeion hansgirgiani Ercegovic 1932: Tab. 11: 215).
Car. all.: Microcoleus lyngbyaceus.
Tipo nomenclaturale: Entophysalidetum deustae Berner 1931
Per quanto riguarda i syntaxa di ordine superiore del Mesolitorale si condivide la proposta di Molinier, precedentemente esposta, che riconosce per ognuno dei sottopiani un ordine ed una alleanza distinti. Le denominazioni proposte da Molinier sono nomina dubia da rigettare in parte a norma dell'art. 2c del C. N.F. (Codice di nomencaltura Fitosociologica), perchè derivati da nomi di animali, ed in tutto a norma dell'art. 38, perchè i nomi dei syntaxa di rango inferiore che li tipificano sono stati considerati nomina dubia. Alcune specie considerate da Molinier come caratteristiche di ordine superiore, sono risultate tali anche a noi e pertanto le abbiamo mantenute nel contingente floristico caratterizzante i vari syntaxa.
Questa considerazione supera la difficoltà, pur presente dal punto di vista formale, data dal fatto che Molinier parla di biocenosi e non di associazioni, come rango inferiore del suo sistema sintassonomico e quindi formalmente si tratta, a norma dell'art. 34, di un rango non previsto dal C. N.F. Noi abbiamo analizzato nelle pagine precedenti le biocenosi di Molinier, in quanto riteniamo, sulla base delle argomentazioni riportarte in un lavoro precedente dell'Autore (Molinier & Picard 1953), che queste biocenosi sono state metodologicamente introdotte con criteri sintassonomci analoghi a quelli che si utilizzano in letteratura per le associazioni ma con liste di specie vegetali completate da liste di specie animali, strettamente legate alle prime e conducenti come le prime vita bentonica. A rafforzare questa nostra opinione interviene lo schema di gerarchizzazione adottato dall'Autore per queste biocenosi quadro, in ranghi di ordine superiore: ordini e alleanze.
In base alle argomentazioni precedenti riteniamo opportuno introdurre nuovi syntaxa di ordine superiore per i due sottopiani del Mesolitorale.
Sottopiano superiore
Ord. Bangietalia atropurpureae Giaccone 1993.
Tipo nomenclaturale: Bangion atropurpureae
Car. ord.: Hapalospongidion macrocarpum.
All. Bangion atropurpureae Giaccone 1993.
Tipo nomenclaturale: Bangietum atropurpureae
Car. all.: Hapalospongidion macrocarpum.
A questi sintaxa di ordini superiore che caratterizzano il sottopiano superiore del mesolitorale in Mediterraneo vanno riferite le associazioni che formano cinture stagionali e popolano orizzonti sovrapposti: Bangietum atropurpureae, Porphyretum leucostictae e Nemalio-Rissoelletum verruculosae.
Sottopiano inferiore
Ord. Ralfsietalia verrucosae Giaccone 1993.
Tipo nomenclaturale: Ralfsion verrucosae
Car. ord.: Ralfsia verrucosa, Nemoderma tingitanum, Gastroclonium clavatum e Corallina elongata.
All. Ralfsion verrucosae Giaccone 1993.
Tipo nomenclaturale: Lithophylletum byssoidis
Car. all.: Ralfsia verrucosa, Nemoderma tingitanum, Gastroclonium clavatum e Corallina elongata.
A questi due syntaxa di ordine superiore che caratterizzano il sottopiano inferiore del mesolitorale in Mediterraneo vanno riferite le seguenti associazioni perennanti, caratterizzate da forte periodismo stagionale:
- Lithophylletum byssoidis (incluse le subassociazioni),
- Ceramio-Corallinetum elongatae,
- Phymatolithetum lenormandii
- e probabilmente anche Fucetum virsoidis (in stazioni dell'Adriatico),
- Enteromorphetum compressae (in biotopi con acque dissalate per presenza di scoli di acque dolci più o meno eutrofiche)
- Polysiphonio-Lithophylletum papillosi.
I tre ordini fitosociologici proposti per il sopralitorale e il mesolitorale, con le rispettiva alleanze e associazioni, si propone di includerle in una unica classe denominata:
Classe Entophysalidetea deustae Giaccone 1993.
Tipo nomenclaturale Lithophylletum byssoidis
Car. cl.: Brachytrichia quojii.
La caratterizzazione ecologica di questa classe è quella riportata nel paragrafo precedente relativo a "Caratteri comuni ai due piani: sopralitorale e mesolitorale"
La vegetazione sommersa
L'associarsi dei vegetali nella zona intertidale è condizionata dalla latitudine, dalla ampiezza e dalla durata delle fasi delle maree, dal moto ondoso e soprattutto dalla temperatura e dall'umidità. Nella zona costantemente coperta dall'acqua i fattori che regolano il formarsi di consorzi vegetali sono la consistenza del substrato, la penetrazione della luce, l'idrodinamismo da moto ondoso o da correnti e, in maniera meno determinante, la temperatura e la salinità. La pressione sembra non influire sulla distribuzione verticale dei vegetali sommersi.
Per i popolamenti epifiti e per quelli che vivono in sottostrato, all'ombra delle forme più sviluppate, hanno un'influenza determinante i fattori biotici e gli scambi energetici. I vegetali condizionati da un'immersione continua popolano i piani infralitorale e circalitorale del sistema fitale. La vegetazione in questi piani, a livello dei grandi gruppi sistematici, ha in tutti i mari del mondo una omogeneità maggiore di quella presente nei piani più superficiali. L'ambiente sommerso, infatti, pur risentendo dei cambiamenti dei fattori ecologici nei limiti del sistema fitale mantiene condizioni abbastanza omogenee a tutte le latitudini, eccettuate quelle estreme dei poli e dell'equatore.
Caratteri strutturali ed ecologici fondamentali del piano infralitorale
Il piano infralitorale è limitato superiormente dal livello occupato da specie vegetali che non possono sopportare emersioni prolungate. Il suo livello inferiore corrisponde alla profondità di compensazione delle Angiosperme e delle alghe fotofile. Naturalmente questa profondità dipende dalla penetrazione della luce che è strettamente legata alla torbidità delle acque. Questo piano corrisponde alle zone megafotica e metriofotica di Ercegovic (in Molinier, 1958). In Mediterraneo questo piano si estende poco oltre i 35 metri di profondità nel bacino occidentale e leggermente più in profondità in quello orientale. Halophila stipulacea è l'Angiosperma che arriva anche a circa 100 metri di profondità nei mari tropicali.
Riedl (1964), a seguito di studi sul mare Adriatico, ipotizza che i movimenti delle acque causati dalle onde possano identificare tre zone:
- dalla superficie fino a circa 3 metri di profondità, la zone di "rottura" delle onde, è caratterizzata da movimenti multidirezionali;
- da 3 a circa 11 metri di profondità i movimenti delle acque sono prevalentemente oscillatori;
- da 11 metri a 20-25 metri (raramente 35m) il flusso dell'acqua ha un unica direzione (moto unidirezionale) che, in assenza di correnti permanenti, dipende da quella delle onde.
La prima e la seconda zona sono separate da un limite la cui profondità corrisponde a 2,5 h, dove h indica la media dell'altezza delle onde; il limite tra la seconda e la terza zona corrisponde ad una profondità il cui valore è compreso tra /2 e , dove corrisponde alla lunghezza dell'onda.
Il piano infralitorale può essere diviso in due sottopiani: sottopiano superiore e sottopiano inferiore, tale diversificazione in sottopiani oltre che dalla luminosità può essere determinata dalla zonazione idrodinamica e dagli alotermoclini.
Sottopiano superiore
Il sottopiano superiore è condizionato da una intensità luminosa compresa tra il 60 e 15% di quella incidente alla superficie, e da idrodinamismo multidirezionale e bidirezionale. Questo sottopiano corrisponde, quindi, alle prime due zone descritte da Riedl. In questo sottopiano si distinguono diversi orizzonti: la frangia infralitorale, ampia pochi decimetri, è la zone di passaggio tra il mesolitorale e l'infralitorale. L'orizzonte alto è caratterizzato da idrodinamismo multidirezionale, mentre l'orizzonte basso dalla prevalenza di idrodinamismo bidirezionale. Le Cistoseire che si affermano in questo sottopiano sono simpodiali in ambienti esposti (Es.: C. amentacea, C. brachycarpa, ecc.) e monopodiali in ambienti riparati (Es.: C. mediterranea, C. elegans, C. sauvageauana, ecc.).
Aggruppamenti fotofili dell'infralitorale superiore
Cystoseiretum strictae Molinier 1958
Le specie caratteristiche dell'associazione sono: Cystoseira amentacea var. stricta (come stricta), Feldmannia paradoxa.
Questa vegetazione è stata descritta per la prima volta da Molinier nel 1958 per le coste di Cap Corse, in stazioni di ambiente battuto. L'Autore, pur descrivendo questo aggruppamento come una Biocenosi, mette in evidenza che la componente faunistica è costituita da specie ubiquiste che incidono limitatamente sulla struttura del popolamento. La componente vegetale è costituita fondamentalmente da Cystoseira amentacea v. stricta e Feldmannia paradoxa che l'Autore indica come specie caratteristiche della biocenosi. L'elenco delle specie caratteristiche è stato integrato successivamente da Boudouresque (1971) e da Giaccone (1971). Questi Autori, oltre alle specie indicate da Molinier, considerano caratteristiche di questo aggruppamento: Laurencia truncata (come L.pinnatifida), Sphacelaria cirrosa (come S.hystrix), Boergeseniella fruticulosa, Herponema valiantei, Bryopsis duplex (come B.disticha), Gelidium pusillum. Tuttavia, Sphacelaria cirrosa e Gelidium pusillum: la prima è stata già indicata da Molinier come caratteristica del Cystoseiretum crinitae, la seconda è caratteristica dell'Enteromorphetum compressae Berner 1931. In Adriatico Cystoseira amentacea v. spicata sostituisce C.amentacea v. stricta, mentre nel bacino orientale domina C.amentacea v. amentacea.
Cystoseiretum strictae subass. Cystoseiretosum tamariscifoliae Giaccone 1972
Le specie differenziali di questa associazione sono: Cystoseira tamariscifolia, Mesophyllum lichenoides, Saccorhiza polyschides, Phyllariopsis brevipes (come Phyllaria reniformis), Asparagopsis armata, Schyzimenia dubyii, Desmarestia ligulata, Halurus equisetifolius.
Questo aggruppamento vegetale, descritto da Giaccone (1972) per lo Stretto di Messina e per il Mare d'Alboran, si afferma in biotopi con acque fresche per fenomeni di upwelling. In queste condizione ambientali C.tamariscifolia vicaria C.amentacea costituendo una subassociazione distinta il cui corteo floristico è simile a quello del Cystoseiretum strictae.
Sulle coste laviche con sorgiva di acqua dolce del Golfo di Napoli e del Golfo di Catania Schyzimenia dubji forma facies differenziali che scendono anche nel Cystoseiretum crinitae, spesso associandosi anche a Cystoseira mediterranea, Grateloupia proteus, e Sebdenia monardiana e a specie dell'ordine Ulvales. Nell'area lusitano-senegalese e, più in generale, sulle coste atlantiche dell'Europa è probabile che l'aggruppamento a C.tamariscifolia e a Saccorhiza polyschides costituisca una associazione autonoma.
Dasycladetum vermicularis Mayhoub 1976
Le specie caretteristiche dell'associazione sono: Dasycladus vermicularis, Polysiphonia ferulacea.
L'aggruppamento si impianta in biotopi calmi o moderatamente battuti, su substrati rocciosi leggermente inclinati, ricoperti da sedimenti sabbiosi, fino a circa 40 cm di profondità. L'Autore considera tra le specie caratteristiche Flabellia petiolata (come Udotea petiolata); questa specie, tipica di associazioni sciafile, si trova insieme a Dasycladus vermicularis probabilmente a causa della ridotta luminosità (dovuta al rimaneggiamento del sedimento e all'esposizione delle stazioni) in cui in cui si sviluppa l'associazione nel Mediterraneo orientale.
Sargassetum vulgaris Mayhoub 1976
Le specie caretteristiche dell'associazione sono: Sargassum vulgare, S.trichocarpum.
Questa associazione, descritta per le coste siriane, sostituisce il Cystoseiretum strictae in stazioni moderatamente battute, raggiungendo il massimo sviluppo in biotopi ombreggiati a circa -2/-3 metri di profondità. S.trichocarpum può formare anche facies estese nel circalitorale del Mare Egeo (Giaccone, 1968-69).
Cystoseiretum crinitae Molinier 1958
Le specie caratteristiche dell'associazione sono: Cystoseira crinita, Sphacelaria cirrosa (come S.hystrix), Stypocaulon scoparium, Cladostephus spongiosus f.verticillatus (come C.verticillatus), Anadyomene stellata.
Molinier nel 1958 descrive la biocenosi quadro Cystoseiretum crinitae per le coste di Cap Corse, questo aggruppamento si trova in ambienti sia di moto battuto che di moto calmo. La componente faunistica, sebbene abbondante, è rappresentata da specie ubiquiste che non incidono sulla struttura dell'aggruppamento, pertanto questa biocenosi può essere considerata un'associazione vegetale. L'Autore indica quali specie caratteristiche: Cystoseira crinita, Sphacelaria cirrosa (come S.hystrix), Stypocaulon scoparium, Cladostephus spongiosus f.verticillatus (come S.verticillatus), Anadyomene stellata.
Successivamente Giaccone (1971) e Boudouresque (1971), trattano questo aggruppamento vegetale riportando delle tabelle di rilievi. In particolare Boudouresque propone di aggiungere alle specie indicate come caratteristiche da Molinier anche: Padina pavonica, Dictyota fasciola (come Dilophus fasciola), Erythrocystis montagnei (come Ricardia montagnei), Liagora viscida, Herposiphonia secunda, Liebmannia leveillei, Dasycladus vermicularis, Cystoseira elegans, C.brachycarpa v.balearica (come C.caespitosa); ma le prime quattro specie, come già proposto da Molinier (1958), sono da considerarsi appartenenti ad unità fitosociologiche di ordine superiore (Alleanza Cystoseirion crinitae), le rimanenti sono caratteristiche di altre Associazioni. Infine C.brachycarpa v. balearica viene considerata come vicariante di crinita. O secondo Pizzuto (1998) come specie elettiva di un aggruppamento vegetale autonomo.
Verlaque (1987) corresse la denominazione di questa associazione in Cystoseiretum balearicae poichè nella stessa località in cui Molinier descrisse il Cystoseiretum crinitae trovò Cystoseira balearica anzichè C.crinita. Questa non è una motivazione legittima a norma del C. N.F., pertanto resta valida la combinazione nomenclaturale proposta originariamente da Molinier (1958).
Cystoseira crinita è la specie più ricca di vicarianti geografiche ed ecologiche già indicate da Giaccone & Bruni (1973): C.abies-marina: regione Macaronesia; C.algeriensis: ambienti caldi e ricchi di sedimentazione; C.barbatula: Mediterraneo orientale (Egeo, Jonio, Canale di Sicilia) che risale anche nella frangia infralitorale; C.brachycarpa v.brachycarpa e v.balearica: Tirreno centro-meridionale; C.corniculata: Adriatico, Egeo e in parte dello Jonio in ambienti con sedimenti; C.hyblaea: Mar Jonio nord-occidentale che risale nella frangia infralitorale; C.elegans: pozze infralitorali ed in baie riparate; C.sedoides: presente in Nord Africa ed a Pantelleria in ambienti poco illuminati; C.susanensis: Tirreno meridionale, Mar Jonio e Mar Africano. Nei settori settentrionali, o in siti con presenza di acque fresche per fenomeni di up-welling, questa specie può essere vicariata anche da C.mediterranea, che risale anche nella frangia.
Cystoseiretum crinitae subass. Alsidietosum helmintochortonis Molinier 1958
La specie differenziale di questa subassociazione è: Alsidium helminthochorton.
Questa subassociazione si afferma in biotopi con forte sedimentazione e idrodinamismo ridotto.
Cystoseiretum crinitae subass. Cystoseiretosum compressae Molinier 1958
La specie differenziale di questa subassociazione è: Cystoseira compressa
Questa specie può risalire anche nel mesolitorale o discendere nel sottopiano inferiore dell'infralitorale.
Molinier descrive questa subassociazione come una facies di impoverimento dell'associazione tipo. Si afferma anche in stazioni relativamente calme e leggermente inquinate.
Cystoseiretum crinitae subass. Stypocauletosum scoparii Boudouresque 1971
La specie differenziale di questa subassociazione è: Stypocaulon scoparium.
Si afferma nei primi metri dell'infralitorale, in biotopi con instabilità ambientale, in particolare, di origine sedimentaria. A questo aggruppamento è possibile ricondurre il popolamento a Padina pavonica e Dictyota fasciola (come Dilophus fasciola) descritto da Mayhoub (1976) per le coste siriane; entrambe queste specie, infatti, sono presenti nei rilievi pubblicati da Boudouresque (1971) con elevati valori di copertura. Stypocaulon scoparium scoparia, Halopteris filicina, Padina pavonica e Dictyota fasciola si ritrovano comunemente in stazioni in cui l'instabilità ambientale, dovuta alla sedimentazione, è la caratteristica predominante.
Cystoseiretum crinitae subass. Halopitetosum incurvae Boudouresque 1971
Le specie differenziali di questa subassociazione sono: Halopitys incurva, Dipterosiphonia rigens.
Si afferma nei primi metri dell'infralitorale, in biotopi con instabilità ambientale, in particolare, di origine fotica.
Cystoseiretum crinitae subass. Gelidietosum spinosi hystricis Marino, Di Martino & Giaccone 1999
La specie caratteristica di questa sub-associazione è: Gelidium spinosum v. hystrix.
Questa sub-associazione si sviluppa sui substrati sub-verticali esposti ad oriente ed insieme alla specie differenziale presenta un ricoprimento elevato di specie caratteristiche di ordine superiore a tallo calcareo (Amphiroa rigida e Jania rubens) Si nota anche la presenza significativa di specie accompagnatrici sempre a tallo calcareo (Haliptilon virgatum, Neogoniolithon brassica-florida, Peyssonnelia polymorpha, Jania adhaerens, Lithophyllum byssoides) oltre che di Fucales (Cystoseira compressa e Sargassum vulgare), di Dictyotales (Dictyopteris polypodioides, Dictyota dichotoma) e di Rhodomelaceae (Boergeseniella fruticulosa, Laurencia majuscula). Questi valori di presenza e di ricoprimento di specie evitate dagli erbivori, denota un forte disturbo, causato da questi organismi sul manto vegetale, caratterizzato dal prevalere di specie non appetibili e non digeribili. Questa sub-associazione ha dunque la composizione di un syntaxon durevole bloccato nell’evoluzione da intensa erbivoria selettiva, in sinergia con l’esposizione e l’inclinazione del substrato. Questo è confermato anche dal fatto che Gelidium spinosum v.hystrix, dove non vi è l’influenza sinergica di questi fattori, si ritrova come specie accompagnatrice con scarsi valori di frequenza e di ricoprimento in tutto l’infralitorale dell’area studiata.
Cystoseiretum barbatae Pignatti 1962
Le specie caratteristiche dell'associazione sono: Cystoseira barbata, C.compressa (come C.abrotanifolia), Halymenia floresia, Gracilaria bursa-pastoris (come G.compressa), Hypnea musciformis, Nemastoma dichotoma, Ceramium diaphanum v. diaphanum, diaphanum v. strictum, C.diaphanum v. lophophorum, Bonnemaisonia asparagoides.
Descritta da Pignatti (1962) per le coste venete, quest'associazione si sviluppa in ambienti caratterizzati da luce e idrodinamismo ridotti. Pignatti inserisce nell'elenco delle specie caratteristiche anche Cladostephus spongiosus f. verticillatus (come C.verticillatus) e Dictyopteris polypodioides; queste specie sono inserite anche tra le specie caratteristiche rispettivamente del Cystoseiretum crinitae Molinier 1958, e del Dictyopteretum polypodioidis Berner 1931.
Trichosoletum myurae Mayhoub 1976
Le specie caratteristiche dell'associazione sono: Liagora C. farinosa, Trichosolen myura, Hydroclathrus clathratus.
L'associazione descritta da Mayhoub (1976) per le coste della Siria si sviluppa in "cuvettes" profonde. Si tratta di un aggruppamento fotofilo di affinità calda e che si sviluppa in ambienti con idrodinamismo ridotto. Tra le specie caratteristiche l'Autore riporta Lophocladia lallemandii e Anadyomene stellata; la prima riteniamo sia ampiamente distribuita, mentre la seconda è stata già indicata da Molinier (1958) come caratteristica del Cystoseiretum C. crinitae.
Herposiphonio-Corallinetum elongatae Ballesteros 1988
Le specie caratteristiche dell'associazione sono: Herposiphonia secunda f. tenella, Corallina elongata.
Questa associazione si afferma in biotopi poco profondi e con elevato idrodinamismo. La specie dominante è Corallina C. elongata, associata a Herposiphonia C. secunda f. tenella, mentre altre specie epifite non assumono un ruolo rilevante. L'Autore descrive quest'aggruppamento per la Costa Brava, ma la sua distribuzione in Mediterraneo è probabilmente più ampia, recentemente è stata riportata per l'Isola di lampedusa. Tuttavia è da rilevare che Corallina elongata è stata indicata come specie caratteristica anche dell'Ordine Ralfsietalia verrucosae Giaccone 1993; si tratta evidentemente di una specie con una ampia distribuzione batimetrica.
Cystoseiretum sauvageauanae (Giaccone 1973 nom. dub.) Giaccone 1994
Le specie caratteristiche dell'associazione sono: Cystoseira sauvageauana, C.schiffneri f. tenuiramosa, Stilophora rhizodes.
Le condizioni ecologiche in cui si afferma questa associazione sono quelle indicata da Giaccone. Come specie caratteristiche vengono indicate: Cystoseira sauvageauana, C.schiffneri f. tenuiramosa, Stilophora rhizodes, mentre Lophocladia lallemandii, presente in tabella tra le specie caratteristiche, viene ritenuta a più larga valenza ecologica. In biotopi del Mediterraneo orientale Cystoseira spinosa v. tenuior vicaria C.sauvageauana. In altri biotopi C. brachycarpa v. brachycarpa può vicariare C.sauvageauana e può discendere anche nell'infralitorale inferiore.
Questa associazione è stata descritta da Giaccone, in Giaccone & Bruni (1973), per il Mediterraneo sud-occidentale e si trova nell'orizzonte basso dell'infralitorale superiore caratterizzato da correnti oscillanti. Per quanto riguarda la tabella completa, l'Autore fa riferimento ad un lavoro precedente, nel quale la tabella IV "popolamenti a Cystoseira C. sauvageauana e brachycarpa", contiene 25 rilievi. Tuttavia nessuno dei rilievi presenti in questa tabella può essere considerato come rilievo tipo. Infatti:
- tra le specie caratteristiche che vengono riportata nel testo e quelle che sono presenti in tabella non c'è corrispondenza;
- le superfici rilevate sono estremamente ampie (secondo la metodologia terrestre) e pertanto sono presenti numerose specie di Cystoseira alcune delle quali caratteristiche di altre Associazioni (C. crinita e C. spinosa);
- il dato quantitativo di C. sauvageauana è dato insieme a quello di C. brachycarpa pertanto l'aggruppamento descritto mostra una copresenza di C. sauvageauana e C. brachycarpa, le cui coperture percentuali sono probabilmente simili. In altre stazioni, invece, C. sauvageauna forma dei popolamenti fisionomicamente omogenei e ben strutturati, in cui C. brachycarpa non è presente.
Pertanto Cystoseiretum sauvageauanae Giaccone 1972 è un nomen dubium secondo l'art. 37 del C. N.F.
Aggruppamenti sciafili dell'Infralitorale superiore
Schotteretum nicaeensis Berner 1931
Le specie caratteristiche dell'associazione sono: Schottera nicaeensis (come Phyllophora palmettoides), Gymnogongrus crenulatus (come G. norvegicus)
"Tipo di associazione di alghe della regione litorale, che si sviluppa in acqua di mare aerata, più o meno inquinata e diluita in maniera intermittente, ma unicamente localizzata nelle stazioni con luce fortemente ridotta a debole profondità".
Rhodymenietum ardissonei Pignatti 1962
Le specie caratteristiche dell'associazione sono: Chondracanthus acicularis, Rhodophyllis divaricata (come Rhodophyllis bifida). L'A. considera Rhodymenia ardissonei, dominante nell'associazione, come specie ad ampia distribuzione batimetrica. Infine vengono individuate due varianti stagionali: una autunnale a Callithamnion corymbosum ed una primaverile a Bryopsis plumosa.
L'ambiente in cui si afferma, oltre che dalla scarsa intensità luminosa in buchi e anfratti dei "murazzi", è caratterizzato da una relativa calma idrodinamica. L'associazione è riportata anche per il Golfo di Trieste. L'ambiente dove sono stati effettuati i rilievi è, al contrario di quello veneziano, di origine naturale, ubicato in superficie ed alquanto esposto alla risacca. La configurazione floristica è di conseguenza affine anche allo Schotteretum nicaeensis Berner 1931.
Pterothamnio-Compsothamnietum thuyoidis Boudouresque, Belsher & Marcot-Coqueugniot 1977.
Le specie caratteristiche dell'associazione sono: Pterothamnion crispum (come Platythamnion plumula v.crispum), Compsothamnion thuyoides.
L'associazione si sviluppa nell'Infralitorale superiore presso la superficie dei piccoli porti non eccessivamente inquinati e sui substrati artificiali dei moli e delle banchine. Gli A.A. la includono nell'ordine Rhodymenietalia Boudouresque 1971, perchè le specie tionitrofile dell'Ordine Ulvetalia Molinier 1958 vi sono scarsamente rappresentate.
Nonostante questo aggruppamento sia stato caratterizzato dagli A.A. con la metodologia dei gruppi ecologici, le informazioni sono riportate in maniera da soddisfare le esigenze fondamentali della metodologia sigmatista e l'associazione quindi risulta validamente descritta.
Sottopiano inferiore
Il sottopiano inferiore è caratterizzato da una luminosità compresa tra il 20 e 1% di quella incidente alla superficie, una temperatura che non supera i 18-20 C e da un idrodinamismo prevalentemente unidirezionale. Le Cistoseire che si affermano in questo sottopiano sono generalmente monopodiali (Es.: C. jabukae, C. spinosa, C. mauritanica); ma in presenza di correnti si affermano anche specie simpodiali (C. brachycarpa v. claudiae, C. corniculata).
Aggruppamenti fotofili dell'Infralitorale inferiore
Cystoseiretum spinosae Giaccone 1973
Le specie caratteristiche dell'associazione sono: Cystoseira spinosa, C. schiffneri v. latiramosa, Valonia macrophysa, Halopteris filicina, Dictyota dichotoma v. intricata (come D. linearis).
Questa associazione descritta da Giaccone, in Giaccone & Bruni (1973), ricopre i substrati duri dell'infralitorale inferiore nelle porzioni meridionali dei bacini occidentali ed orientali del Mediterraneo, caratterizzati da idrodinamismo ridotto e di tipo fluente. Per quanto riguarda la tabella completa, l'Autore fa riferimento ad un lavoro precedente, Giaccone 1971, nel quale la tabella V "aspetti della biocenosi coralligena" contiene 24 rilievi di cui 13 relativi a questo aggruppamento. Ma, non tutte le specie che vengono riportata nel testo come caratteristiche sono presenti in tabella; pertanto riteniamo specie caratteristiche dell'agguppamento solo quelle che sono presenti sia nella tabella che nel testo: Cystoseira spinosa, C. schiffneri v. latiramosa, Valonia macrophysa, Halopteris filicina, Dictyota dichotoma v. intricata (come D. linearis). Dall'analisi della tabella il popolamento non appare omogeneo dato che C. spinosa ha scarsi valori di copertura. Questa specie in altre stazioni del Mediterraneo raggiunge coperture elevate costituendo popolamenti omogenei e ben strutturati. In Adriatico Cystoseira spinosa è vicariata da C. corniculata in ambienti con forte sedimentazione, mentre nel Tirreno, in ambienti con sedimentazione grossolana e in presenza di correnti è presente C. jabuka. Il limite inferiore dell'infralitorale, marcato da questa vegetazione a Cystoseire, può estendersi anche al di sotto del limite inferiore delineato dalle Angiosperme marine.
Aggruppamenti sciafili dell'Infralitorale inferiore.
Flabellio-Peyssonnelietum squamariae Molinier 1958.
Le specie caratteristiche dell'associazione sono: Peyssonnelia squamaria, Flabellia petiolata (come Udotea petiolata), Osmundaria volubilis (come Vidalia volubilis).
Molinier (1958) descrisse l'aggruppamento vegetale sciafilo che si sviluppa sui rizomi e più in generale nel sottostrato della prateria a Posidonia oceanica. Egli la denominò biocenosi sciafila dei rizomi, anche se la componente faunistica riportata nei rilievi non è nè determinante nè caratterizzante del contingente fondamentale, in effetti l'A. la denomina con terminologia fitosociologica Udoteo-Peyssonnelietum Molinier 1958. L'A. inquadra questa associazione, insieme al Posidonietum oceanicae Molinier 1958, nella Classe Zosteretea Pignatti 1953.
Secondo Pignatti (1953) questa classe raggruppa soltanto la vegetazione marina ad Angiosperme. Boudouresque (1971) correttamente include questa associazione nell'Ordine Rhodymenietalia Boudouresque 1971 e di fatto emenda l'associazione sciafila, anche se non lo dichiara formalmente, sia nella componente floristica caratterizzante (vedi tab. X e XI dell'A., 1971), che nell'inquadramento ambientale (biotopi sciafili calmi dell'Infralitorale inferiore). Pérès e Picard (1964) ed altri ricercatori dopo di loro considerano questa associazione la componente vegetale di una complessa formazione sciafila denominata "Precoralligeno".
Anche se i rilevamenti successivi a quelli effettuati da Molinier (1958) sono realizzati secondo l'estensione ambientale introdotta da Boudouresque (1971), e che noi condividiamo, riteniamo che l'associazione introdotta da Molinier resti sufficientemente definita e caratterizzata floristicamente nella sua presentazione originale.
Osmundarietosum volubilis Serio & Pizzuto 1992.
La specie differenziale della subassociazione è: Osmundaria volubilis.
Questa subassociazione del Flabellio-Peyssonnelietum squamariae Molinier 1958, è stata descritta per l'Infralitorale della Sicilia orientale in ambienti scarsamenti illuminati e soggetti ad intensa sedimentazione. È da precisare che in questi biotopi l'Infralitorale ha il suo limite inferiore intorno ai -20 metri.
Molti altri ricercatori in precedenza avevano segnalato in differenti settori biogeografici del Mediterraneo questo aggruppamento. La specie guida di questa subassociazione può estendersi anche nel Circalitorale, sia su substrato duro che nella biocenosi del detritico costiero.
Halymenietum floresiae Giaccone & Pignatti 1967.
Le specie caratteristiche dell'associazione sono: Halymenia floresia, Boergeseniella fruticulosa (come Polysiphonia fruticulosa), Halymenia dichotoma (come Chrysimenia dichotoma), Cladophora prolifera, Scinaia furcellata, Sphaerococcus coronopifolius, Chrysimenia ventricosa, Halarachnion ligulatum, Thuretella schousboei, Alsidium corallinum.
Gli A.A. includono tra le caratteristiche Zanardinia prototypus, ma Giaccone considera questa specie caratteristica dell'alleanza Sargassion hornschuchii Giaccone 1973. Si tratta in effetti di una specie emisciafila diffusa nell'Infralitorale, ma che può penetrare anche in tutto il Circalitorale. L'associazione si sviluppa nell'Infralitorale in biotopi caratterizzati da risorgive marine di acque dolci e sopporta anche acque moderatamente eutrofiche. L'Infralitorale nel Golfo di Trieste ha il suo limite inferiore intorno a 7 - 10 metri di profondità. L'associazione è descritta per il Golfo di Trieste, ma è possibile riscontrarla anche in altri biotopi del Mediterraneo.
Rhodymenio-Codietum vermilarae Ballesteros 1989.
Le specie caratteristiche dell'associazione sono: Codium vermilara, Aglaothamnion (come Callithamnion) tripinnatum, Spermothamnion flabellatum.
L'A. descrisse questa associazione per la Costa Brava, ma la ritiene diffusa in tutta la costa catalana. Si afferma in ambienti calmi dell'Infralitorale inferiore, dove sostituisce il Cystoseiretum spinosae Giaccone 1973. Si sviluppa principalmente nel periodo primaverile-estivo tra gli 8 ed i 20 m. di profondità, in biotopi caratterizzati da scarsa intensità luminosa (12-2% di quella misurata in superficie) e con una relativa ricchezza di nutrienti azotati. L'aggruppamento è descritto dall'A. con il metodo dei gruppi ecologici, ma è riconducibile alle esigenze essenziali del metodo sigmatista e delle norme del C.N.F. Riteniamo di interpretare correttamente il testo dell'A. indicando più formalmente le specie caratteristiche sopra indicate. Tra le specie che presentano una netta dominanza qualitativa e quantitativa l'A. riporta Rhodymenia ardissonei e Mesophyllum lichenoides, dotate di più larga ripartizione batimetrica. L'A., infine, considera strettamente legate al contingente caratterizzante le specie a distribuzione boreale: Anthithamnionella spirographidis e Audouinella codii.
Caratteri strutturali ed ecologici fondamentali del Circalitorale
Si estende al di sotto del limite inferiore dell'infralitorale, fino alla profondità massima alla quale le alghe multicellulari possono esplicare la fotosintesi. Questo piano, che corrisponde alla zona oligofotica di Ercegovic, in Mediterraneo, su substrato duro, è condizionato da una intensità luminosa compresa tra 0,9% e 0,01% di quella misurata in superficie. La temperatura oscilla tra 18°C e 13,5° L'idrodinamismo è dato da correnti di gradiente termoalino e di dislivello tra bacini, potenziato da correnti di stretto in biotopi separati da strozzature geomorfologiche. Il limite inferiore del manto vegetale generalmente si interrompe tra 120 e 150 metri di profondità. Specie algali pluricellulari sono state raccolte fino a 200 metri di profondità ed in tutta la piattaforma continentale, ma si tratta di specie non strutturate in associazioni vegetali. Comprende due sottopiani: nel sottopiano superiore che finisce mediamente intorno a 80 metri di profondità è possibile ancora trovare una vegetazione stratificata con in strato elevato Fucales (Cystoseira e Sargassum), Laminariales, Desmarestiales e Sporochnales ed in sottostrato Rodoficee molli ed incrostanti insieme a poche Cloroficee (Palmophyllum, Codium e altri generi). Nel sottopiano inferiore tra 80-130 metri di profondità si ritrova una importante copertura vegetale ad opera di Melobesie libere e di specie algali aggreganti e reptanti. A profondità maggiore le forme vegetali presenti spesso sono cadute dal livello superiore e sopravvivono stentatamente. Pertanto la vegetazione fotofila che si afferma in questo piano è rappresentata da Aggruppamenti vegetali in cui è presente uno strato elevato ben sviluppato.
Aggruppamenti fotofili del Circalitorale
Cystoseiretum zosteroidis Giaccone 1973
Le specie caratteristiche dell'associazione sono: Cystoseira zosteroides, Arthrocladia villosa, Sporochnus pedunculatus, Polysiphonia foeniculacea.
Questa associazione è stata descritta da Giaccone, in Giaccone & Bruni (1973), per biotopi caratterizzati da correnti di fondo a direzione costante, da un'intensità luminosa compresa tra 0,9% e 0,1% di quella incidente sulla superficie e da una temperatura che oscilla stagionalmente tra 16 °C e 14 ° Per quanto riguarda la tabella completa, l'Autore fa riferimento a tabelle pubblicate in lavori precedenti (Giaccone 1965, 1967, 1969, 1971). Tra queste riteniamo che la più completa sia quella in Giaccone 1971: tabella V "aspetti della biocenosi coralligena" con 24 rilievi, di cui 11 relativi a questo aggruppamento. In questa tabella, tuttavia, non sono presenti tutte le specie che vengono indicate nel testo come caratteristiche; pertanto riteniamo tali solo quelle presenti sia nella tabella che nel testo: Cystoseira zosteroides, Arthrocladia villosa, Sporochnus pedunculatus, Polysiphonia foeniculacea. E da notare che i rilievi sono stati effettuati a notevole profondità (da -40 a -80 m), ciò potrebbe spiegare la scarsa copertura della specie guida (da 1 a 20%) e l'abbondanza quantitativa e qualitativa di specie sciafile.
Questo aggruppamento è stato descritto anche per le coste catalane; in quest'area, il Cystoseiretum zosteroidis è presente ad un livello batimetrico superiore (-16/-18 m), la specie guida presenta maggiori valori di copertura. Osservazioni simili sono riportate per le coste orientale della Sicilia.
Nel Mare Egeo e in maniera impoverita anche nel Mar Jonio (orientale) e nel Mare Adriatico (medio e basso) C. zosteroides è vicariata da C. corniculata che si afferma in ambienti con caratteristiche ecologiche simili a quelle in cui si sviluppa il Cystoseiretum zosteroidis.
Cystoseiretum zosteroidis subass. Laminarietosum rodriguezii Giaccone 1973
La specie differenziale è: Laminaria rodriguezii.
Questa subassociazione descritta in Giaccone e Bruni (1973) si afferma in biotopi al di sotto dei 40 metri di profondità con scarsa luminosità e in presenza di correnti di fondo di direzione costante. Il corteo floristico di questa subassociazione è uguale a quello del Cystoseiretum zosteroidis, ma vi è una maggiore rappresentanza di specie sciafile e reofile.
Cystoseiretum usneoidis Giaccone 1972
Le specie carratteristiche diell'associazione sono: Cystoseira usneoides, Laminaria ochroleuca, Phyllariopsis purpurascens (come Phyllaria purpurascens), Ulva olivascens, Callophyllis laciniata, Phyllophora heredia.
Questa associazione descritta da Giaccone (1972), si afferma in aree limitate del Mediterraneo (Mare di Alboran, Stretto di Gibilterra e Stretto di Messina). L'area geografica dell'Atlantico orientale ospita la stessa associazione, ma ad una profondità inferiore (10-15 m) di quella del Mediterraneo. Le condizioni ambientali che determinano l'affermarsi dell'associazione sono: correnti intense di tipo pulsante; l'esistenza di una accentuata omotermia; ricchezza di sali nutritivi. L'intensità luminosa è compresa da 1 e 0,05% di quella incidente sulla superficie.
Cystoseiretum usneoidis subass. Laminarietosum ochroleucae Giaccone 1994.
Le specie differenziali sono: Laminaria ochroleuca, Lithothamnion philippi.
Quest'aggruppamento si afferma in biotopi (Stretto di Messina, Mare di Alboran) più profondi (50-70 m di profondità) caratterizzati dalla presenza di forti correnti pulsanti. In queste condizioni Cystoseira usneoides assume un ruolo secondario mentre L. ochroleuca costituisce l'elemento dominante e a volte esclusivo dello strato elevato, con il sottostrato prevalentemente costituito da Lithothamnion philippi. Il popolamento a L. ochroleuca, descritto come facies, sembra individuare una subassociazione autonoma che viene descritta considerando olotipo il rilievo 13/29 della tabella II in Giaccone 1972. In questa tabella, che riporta i rilievi del Cystoseiretum usneoidis, sono presenti alcuni rilievi in cui L. ochroleuca assume significativi valori di copertura.
Cystoseiretum dubiae Furnari, Scammacca, Cormaci e Battiato 1977
Le specie caratteristiche dell'associazione sono: Cystoseira dubia, Nitophyllum tristromaticum, Kallymenia patens.
Questo aggruppamento è stato descritto, come "popolamento a C. dubia", per le coste della Sicilia orientale (Isola Lachea, Catania), in presenza di correnti periodiche e unidirezionali e forte sedimentazione. Il Cystoseiretum dubiae si sviluppa intorno ai 40 metri di profondità ed è caratterizzato da discreta copertura della specie guida, soprattutto nei mesi estivi, e da Kallymenia patens e Nitophyllum tristromaticum che vengono pertanto considerate specie caratteristiche. Gli Autori riportano una tabella con 113 rilievi, quelli relativi al Cystoseiretum dubiae vanno dal 103 al 113. C. dubia è stata raccolta fino a circa 200 metri di profondità nel Mediterraneo oriantale e oltre 100 metri di profondità nel Mediterraneo occidentale.
Aggruppamenti sciafili del Circalitorale.
Lithophyllo-Halimedetum tunae Giaccone 1965
Le specie caratteristiche dell'associazione sono: Lithophyllum stictaeforme (come Pseudolithophyllum expansum), Halimeda tuna (come Halimeda platidysca).
L'A. indica fra le specie caratteristiche anche Flabellia petiolata (come Udotea petiolata), già inserita da Molinier (1958) tra le specie caratteristiche del Flabellio-Peyssonnelietum squamariae Molinier 1958. Lo stesso vale per Codium bursa considerata specie caratteristica dell'alleanza Sargassion hornschuchii Giaccone 1973. L'A. riporta una specie differenziale: Peyssonelia polymorpha, è probabile che in questo taxon sia inclusa anche Peyssonnelia rosa-marina, a quel tempo ancora non separata tassonomicamente da P. polymorpha.
Anche se tutte le specie citate hanno un'ampia ripartizione nei biotopi sciafili, formano un'associazione nel Circalitorale superiore, dove Halimeda tuna forma prati estesi. Questa associazione può risalire anche nell'Infralitorale inferiore. L'ambiente di elezione è caratterizzato da debole torbidità, presenza di idrodinamismo unidirezionale costante e con apporti di acque fresche dal fondo. La componente animale bentonica manifesta queste condizioni con il prevalere di filtratori. Ballesteros (1991) ha realizzato uno studio approfondito sulla ricchezza floristica, sulla biomassa e sull'ecologia di questa associazione lungo i fondali della Costa dei Pirenei davanti a Tossa del Mar. Questo A. evidenzia una vicarianza di Mesophyllum lichenoides, che prevale nettamente su Lithophyllum stictaeforme, in condizione di maggiore intensità di energia luminosa. Questo fenomeno è stato riscontrato anche in Sicilia sud-orientale.
Rodriguezelletum strafforellii Augier & Boudouresque 1975
Le specie caratteristiche dell'associazione sono: Rodriguezella strafforellii, Rodriguezella pinnata, Spermothamnion johannis, Ceramium bertholdii, Sphacelaria plumula, Blastophysa rhizopus, Polysiphonia subulifera.
Gli A.A. citano anche i rilievi riportati in Boudouresque (1973), come caratterizzanti la Sub-biocenosi del Coralligeno circalitorale denominata in quel lavoro Peyssonnelietum Rodriguezelletosum Boudouresque 1973 e dipendente dal Peyssonnelietum rubrae Boudouresque 1973. Essi introducono il Rodriguezelletum strafforellii come una biocenosi dei substrati duri di ambienti relativamente calmi del Circalitorale e considerano il contingente fondamentale condizionato in parte da cianofilia obbligatoria. Utilizzano la metodologia dei gruppi ecologici ed indicano tra il contingente caratteristico altre specie non riportate nelle tabelle di riferimento. Nonostante queste carenze ci sembra che ci siano gli elementi essenziali per considerare validamente descritta questa associazione secondo le esigenze del metodo sigmatista e le norme del C.N.F. Questa associazione impoverita si ritrova anche nel sottostrato del Cystoseiretum zosteroidis Giaccone 1973.
Phymatholitho-Lithothamnietum corallioidis Giaccone 1965.
Le specie caratteristiche dell'associazione sono: Phymatholithon calcareum (come Lithothamnion calcareum), Lithothamnion corallioides (come Lithophyllum solutum). L'A. include tra le specie caratteristiche anche Arthrocladia villosa, epibionte sulle Rodoliti, che poi trasferisce tra il contingente caratteristico del Cystoseiretum zosteroidis Giaccone 1973.
Secondo Pérès e Picard (1964) questo aggruppamento è una facies della Biocenosi delle sabbie grossolane e ghiaie fini in correnti di fondo del Detritico costiero, comunemente conosciuta in Atlantico con il nome di Mäerl. In Mediterraneo l'associazione, detta comunemente a Melobesie libere, si sviluppa nel Circalitorale, ma può risalire anche nell'Infralitorale. Ha il suo ambiente favorevole alla base delle falesie in biotopi con correnti di fondo di tipo laminare ad andamento regolare, a profondità di 30-90 metri nel Mediterraneo occidentale e di 80-120 metri nei settori più meridionali ed orientali. Cambiando l'idrodinamismo, la granulometria e l'intensità luminosa si possono avere alcune facies differenziali e/o vicarianze a:
- 1) Peyssonnelia rosa-marina (fango fluido e mobile in correnti turbolenti associate ad eventi di tempesta);
- 2) Lithophyllum racemus (sabbie grossolane e ghiaie fini in correnti turbolenti di fondo);
- 3) Lithothamnion valens (facies a "prâlines" in correnti di fondo laminari e con occasionali picchi di correnti forti);
- 4) Neogoniolithon brassica-florida (come N. notarisii e N. mamillosum), Mesophyllum lichenoides, Lithothamnium fruticulosum auct! (correnti oscillanti e pulsanti in ambienti anche infralitorali con bocche di collegamento con il mare aperto e nei canaloni di intermattes delle praterie a Posidonia oceanica: vedi anche Calvo et al., 1982).
Gli epibionti vegetali sono documentati per il Mediterraneo occidentale da Augier e Boudouresque (1978) e per quello orientale da Giaccone (1968). Formazioni di Rodoliti si ritrovano fossili sia nelle rocce sedimentarie del Mesozoico, sia soprattutto in quelle del Terziario e del Quaternario. Questa associazione è la più estesa formazione vegetale di substrato mobile del Circalitorale in Mediterraneo, paragonabile per importanza alle praterie ad Angiosperme marine dell'Infralitorale.
Syntaxa di ordine superiore: substrato roccioso o duro
La vegetazione fotofila sui substrati rocciosi del Mediterraneo è stata inquadrata da Molinier (1958) in un ordine Cystoseiretalia, in una alleanza Cystoseirion e in due associazioni Cystoseiretum strictae e Cystoseiretum crinitae. L'ordine Cystoseiretalia includeva esclusivamente gli aggruppamenti dell' infralitorale superiore.
Giaccone (1965) introdusse la classe Cystoseiretea Giaccone 1965 che comprende i popolamenti bentonici su substrato roccioso o su sabbia grossolana mista a ghiaia, caratterizzati da una vegetazione stratificata con alghe brune generalmente in strato elevato. Successivamente Giaccone, in Giaccone & Bruni (1973), ha ampliato le caratteristiche ecologiche dell'ordine Cystoseiretalia Molinier 1958 includendo in esso tutte le associazioni fotofile dell'infralitorale e del circalitorale. Inoltre Giaccone propose due nuove Alleanze: Sargassion vulgaris Giaccone 1973 e Sargassion hornschuchii Giaccone 1973. Nella prima di queste due Alleanze incluse, oltre alle due Associazioni di Molinier, anche il Cystoseiretum sauvageauanae Giaccone 1973 e nella seconda Alleanza il Cystoseiretum spinosae Giaccone 1973, il Cystoseiretum zosteroidis Giaccone 1973 ed il Cystoseiretum usneoidis Giaccone 1972. Qui riteniamo valido il Cystoseirion crinitae Molinier 1958, e quindi il Cystoseiretum sauvageauanae viene incluso in questa Alleanza. Il Sargassion vulgaris diviene dunque sinonimo posteriore dell'allenza di Molinier.
Pertanto, la vegetazione marina dell'infralitorale e del circalitorale su substrato roccioso è inquadrata fitosociologicamente secondo il seguente schema:
- Classe Cystoseiretea Giaccone 1965
Tipo nomenclaturale: Cystoseiretalia Molinier emend. Giaccone 1994.
Car. Classe: Jania rubens, Lithophyllum incrustans, Padina pavonica, Dasycladus vermicularis, Dictyota fasciola v. repens, Laurencia obtusa, Acetabularia acetabulum (come A. mediterranea), Pseudolithoderma adriaticum (come Lithoderma adriaticum), Erythrocystis montagnei (come Ricardia montagnei), Amphiroa rigida, Liagora viscida.- Ordine Cystoseiretalia Molinier 1958 emend. Giaccone 1994
Tipo nomenclaturale: Cystoseirion crinitae Molinier 1958.
Car. Ord.: Jania rubens, Lithophyllum incrustans, Padina pavonica, Dasycladus vermicularis, Dictyota fasciola v. repens, Laurencia obtusa, Acetabularia acetabul um (come A. mediterranea), Pseudolithoderma adriaticum (come Lithoderma adriaticum), Erythrocystis montagnei (come Ricardia montagnei), Amphiroa rigida, Liagora viscida.
Giaccone (1973), inserendo nell'ordine Cystoseiretalia Molinier 1958 gli aggruppamenti dell'infralitorale inferiore, ha di fatto emendato questo Syntaxon, cambiandone i caratteri ecologici.- Alleanza Cystoseirion crinitae Molinier 1958
Den orig.: Cystoseirion Molinier 1958.
Tipo nomenclaturale: Cystoseiretum crinitae Molinier 1958.
Car. All.: Jania rubens, Lithophyllum incrustans, Padina pavonica, Dasycladus vermicularis, Dictyota fasciola v. repens, Laurencia obtusa, Acetabularia acetabulum (come A. mediterranea), Pseudolithoderma adriaticum (come Lithoderma adriaticum), Erythrocystis montagnei (come Ricardia montagnei), Amphiroa rigida, Liagora viscida.
Quest'Alleanza è caratterizzata da un idrodinamismo di tipo multidirezionale (fino ad una profondità di circa 2,5 h) e bidirezionale pendolare con forti oscillazioni; l'intensità luminosa compresa tra il 60% e 15% di quella incidente sulla superficie dell'acqua (Giaccone & Bruni, 1973).
Comprende le Associazioni vegetali dell'infralitorale superiore:- Cystoseiretum strictae
- Dasycladetum vermicularis
- Sargassetum vulgaris
- Cystoseiretum crinitae
- Cystoseiretum sauvageauanae
- Cystoseiretum barbatae
- Trichosoletum myurae
- Herposiphonio-Corallinetum elongatae ecc.
- Alleanza Sargassion hornschuchii Giaccone 1973
Tipo nomenclaturale: Cystoseiretum spinosae Giaccone 1973.
Specie caratteristiche: Sargassum hornschuchii, Codium bursa, Spatoglossum solieri, Zanardinia prototypus, Zonaria tournefortii.
È caratterizzata da idrodinamismo unidirezionale fluente. L'intensità luminosa è compresa tra il 20% e 0,01% di quella incidente alla superficie, la temperatura oscilla mediamente tra 14°C e 16°.
L'Alleanza comprende i Cistoseireti dell'infralitorale inferiore e quelli del circalitorale:- Cystoseiretum spinosae
- Cystoseiretum zosteroidis
- Cystoseiretum usneoidis
- Cystoseiretum dubiae
- Alleanza Cystoseirion crinitae Molinier 1958
- Ordine Cystoseiretalia Molinier 1958 emend. Giaccone 1994
Syntaxa di ordine superiore della vegetazione sciafila
La vegetazione sciafila del Mediterraneo nel suo complesso è stata inquadrata storicamente da Giaccone (1965), che distinse due Classi: una per l'Infralitorale ed una per il Circalitorale.
Nella Classe Cystoseiretea Giaccone 1965, nell'Ordine Cystoseiretalia Molinier 1958 e nell'Alleanza Cystoseirion crinitae Molinier 1958, Giaccone incluse lo Schotteretum nicaeensis Berner 1931 (come Petroglosso-Plocamietum Molinier 1958) ed il Flabellio-Peyssonnelietum squamariae Molinier 1958 (come Udoteo-Peyssonnelietum Molinier 1958).
Nella Classe Lithophylletea Giaccone 1965, nell'Ordine Lithophylletalia Giaccone 1965 e nell'Alleanza Lithophyllion stictaeformis Giaccone 1965 (come Lithophyllion), Giaccone incluse il Lithophyllo-Halimedetum tunae Giaccone 1965 (come Pseudolithophyllo-Halimedetum platidyscae Giaccone 1965) ed il Phymatholitho-Lithothamnietum corallioidis Giaccone 1965 (come Lithothamnio-Lithophylletum soluti Giaccone 1965).
Boudouresque (1971: 117-118), introdusse l'Ordine Rhodymenietalia Boudouresque 1971, includendovi le due Associazioni infralitorali create da Molinier (1958). Egli però non tenne presente che Giaccone (1965) aveva introdotto una gerarchizzazione sintassonomica differenziata per la vegetazione sciafila distribuita nei due piani del sistema fitale: Infralitorale e Circalitorale. Di consequenza nel proctologo scrisse che l'ordine da lui introdotto "raggruppa l'insieme dei popolamenti sciafili dell'Infralitorale e del Circalitorale". Di fatto però le due Associazioni che l'A. incluse in quest'Ordine sono, per sua esplicita affermazione e per la descrizione dei loro Autori, a ripartizione solo infralitorale. Di consequenza è stata emendata formalmente la delimitazione batimetrica di questo Ordine limitandola all'Infralitorale. Inoltre si indica come tipo nomenclaturale, non indicato dal suo A., l'Alleanza Peyssonnelion squamariae (come Peyssonnelion) Augier e Boudouresque 1975 e per quest'ultimo syntaxon come tipo nomenclaturale si considera l'associazione Flabellio-Peyssonnelietum squamariae Molinier 1958.
L'altra alleanza Schotterion nicaeensis (come Petroglossion) Boudouresque e Cinelli 1971, era stata introdotta per le due associazioni sciafile di ambiente battuto dell'Infralitorale superiore, da noi considerate nelle pagine precedenti come nomina dubia e ricondotte all'associazione Schotteretum nicaeensis Berner 1931. Pertanto anche questa alleanza risulta emendata, con l'inclusione di altre associazioni che preciseremo più avanti e con l'introduzione del suo tipo nomenclaturale nello Schotteretum nicaeensis Berner 1931.
Il Peyssonnelion squamariae Augier & Boudouresque 1975 è stato introdotto dagli A.A. per raggruppare due associazioni, una diffusa nell'Infralitorale inferiore (Udoteo-Aglaothamnietum tripinnati Augier & Boudouresque, 1975, da noi considerata un sinonimo posteriore del Flabellio-Peyssonnelietum squamariae Molinier 1958) e l'altra nel Circalitorale (Rodriguezelletum strafforellii Augier & Boudouresque 1975).
Gli A.A. nel delimitare l'ambito dell'alleanza non hanno tenuto conto, che per gli aggruppamenti sciafili del Circalitorale Giaccone (1965) aveva creato l'alleanza Lithophyllion stictaeformis (come Lithophyllion) Giaccone 1965. Pertanto si ritiene necessario emendare l'alleanza di Augier & Boudouresque (1975) riportandola nell'ambito dell'Infralitorale inferiore escludendovi il Rodriguezelletum strafforellii ed includendovi altre Associazioni come preciseremo più avanti.
Con riferimento a quanto esposto e documentato in questo paragrafo ed in quelli precedenti proponiamo per la gerarchizzazione della vegetazione sciafila dell'Infralitorale e del Circalitorale del Mediterraneo il seguente schema:
Classe Lithophylletea Giaccone 1965 emend. Giaccone 1994.
Tipo nomenclaturale: Ordine Lithophylletalia Giaccone 1965.
Car. Classe: Lithophyllum stictaeforme, Peyssonnelia rubra, Peyssonnelia inamoena (al tempo non distinta da P. rubra).
La Classe comprende la vegetazione sciafila e di sottostrato dell'Infralitorale e del Circalitorale sia di substrato duro che di substrato mobile, in differenti gradienti di luce, temperatura, idrodinamismo, nutrienti. Nell'emendamento si precisa il tipo nomenclaturale, si indicano le specie caratteristiche e vi si include l'Ordine Rhodymenietalia. Pertanto la Classe emendata comprende i due ordini seguenti:
I. Ordine Rhodymenietalia Boudouresque 1971 emend. Giaccone 1994.
Tipo nomenclaturale: Alleanza Peyssonnelion squamariae Augier & Boudouresque 1975 emend. Giaccone.
Car. Ord.: Phyllophora crispa, Valonia macrophysa, Botryocladia botryoides, Mesophyllum lichenoides, Cutleria chilosa, Eupogodon planus, Nereia filiformis.
L'Ordine comprende gli Aggruppamenti sciafili dell'Infralitorale in differenti condizioni idrodinamiche ed inquadrati nelle due Alleanze di seguito caratterizzate. L'emendamento consiste nel restringerne i limiti batimetrici all'Infralitorale, quelli sintassonomici alle Associazioni incluse nelle due Alleanze sotto delimitate e nell'indicare il tipo nomenclaturale e le specie caratteristiche secondo le norme del C.N.F.
I,1 - Alleanza Schotterion nicaeensis Boudouresque & Cinelli 1971 emend. Giaccone 1994.
Den Orig.: Petroglossion Boudouresque & Cinelli 1971
Tipo nomenclaturale: Schotteretum nicaeensis Berner 1931.
Car. All.: Lomentaria articulata, Cruoria cruoriaeformis (come Cruoria purpurea e Contarinia cruoriaeformis), Lomentaria clavellosa (come Chylocladia clavellosa), Halurus flosculosus.
L'Alleanza comprende le Associazioni sciafile dell'Infralitorale in differenti condizioni idrodinamiche di tipo sia multidirezionale che bidirezionale ed in acque sia oligotrofiche che eutrofiche. Le specie indicate come caratteristiche sono incluse nel tipo nomenclaturale su indicato. Comprende le Associazioni sciafile di seguito riportate:
- Schotteretum nicaeensis Berner 1931
- Rhodymenietum ardissonei Pignatti 1962
- Pterothamnio-Compsothamnietum thuyoidis Boudouresque, Belsher & Marcot-Coqueugniot 1997.
I, 2 Alleanza: Peyssonnelion squamariae Augier & Boudouresque 1975 emend. Giaccone 1994.
Den. orig.: Peyssonnelion Augier & Boudouresque 1975.
Tipo nomenclaturale: Flabellio-Peyssonnelietum squamariae Molinier 1958.
Car. All.: le stesse specie indicate sopra come caratteristiche dell'ordine Rhodymenietalia.
Le specie indicate come caratteristiche sono comuni all'ordine Rhodymenietalia e si ritrovano nel tipo nomenclaturale sopra indicato.
L'alleanza comprende le associazioni sciafile dell'Infralitorale in condizioni di idrodinamismo unidirezionale più o meno oscillante, originate dal moto ondoso. La temperatura è ancora sensibilmente influenzata dal periodismo stagionale. Alcune delle Associazioni elencate possono ritrovarsi impoverite nel sottostrato di Angiosperme e Fucoficee. Vi includiamo le seguenti Associazioni sciafile:
- Flabellio-Peyssonnelietum squamariae Molinier 1958
- Osmundarietosum volubilis Serio & Pizzuto 1992
- Halymenietum floresiae Giaccone & Pignatti 1967
- Rhodymenio-Codietum vermilarae Ballesteros 1989
II. Ordine Lithophylletalia Giaccone 1965.
Tipo nomenclaturale: Alleanza Lithophyllion stictaeformis Giaccone 1965.
Car. Ord.: Halimeda tuna, Peyssonnelia polymorpha, P. rosa-marina, Rhodymenia pseudopalmata, Polysiphonia sanguinea.
L'Ordine comprende le associazioni sciafile del Circalitorale sia su substrato duro che mobile, in ambienti calmi o in presenza di correnti di fondo laminari e turbolente di varia intensità. L'Autore ne completa qui la caratterizzazione indicando formalmente il tipo nomenclaturale e le specie caratteristiche. Vi è compresa la seguente Alleanza:
II,1 - Alleanza: Lithophyllion stictaeformis Giaccone 1965. Den. orig.: Lithophyllion Giaccone 1965. Tipo nomenclaturale: Lithophyllo-Halimedetum tunae Giaccone 1965. Car. All.: le specie caratteristiche sono comuni all'Ordine Lithophylletalia. L'Alleanza comprende le Associazioni sciafile del Circalitorale di substrato duro e mobile in condizioni di luminosità prevalentemente diffusa e di temperatura costante o scarsamente influenzata dal periodismo stagionale. L'ambiente può essere calmo o interessato da idrodinamismo laminare o turbolento. Vi includiamo le seguenti Associazioni:
- Lithophyllo-Halimedetum tunae Giaccone 1965;
- Phymatholitho-Lithothamnietum corallioidis Giaccone 1965;
- Rodriguezelletum strafforellii Augier & Boudouresque 1975.
L'inclusione di quest'ultima associazione, è da considerare come emendamento di Giaccone all'originario inquadramento di Augier & Boudouresque (1975) nell'Ordine Rhodymenietalia e nell'Alleanza Peyssonnelion squamariae.
La vegetazione ad Angiosperme
Il substrato (dott. Marcello Catra)
Il termine substrato viene qui utilizzato in riferimento a quelle strutture con le quali gli organismi mantengono un rapporto temporaneo o permanente di contatto. A differenza dei fattori ambientali che regolano la vita nell’ambiente marino quali luce, temperatura, salinità ecc., risulta difficoltoso definire in termini chimico-fisici le proprietà del substrato Il substrato non condiziona direttamente la crescita e lo sviluppo delle alghe bentoniche, ma ha sicuramente una funzione di supporto permettendo alle alghe di fissarsi e di attingere alle sostanze nutritive del mezzo circostante. Soltanto nel caso delle alghe rizofite, delle alghe endolitiche e delle Angiosperme marine il substrato rappresenta almeno in parte una risorsa dei nutrienti richiesti. Quindi per la maggior parte dei vegetali marini risulta indifferente la composizione chimica del substrato mentre sono condizionati dalle caratteristiche tessiturali di quest’ultimo. Infatti alcune specie preferiscono ancorarsi su roccia e/o substrati duri altre crescono esclusivamente su sedimenti sciolti (± substrati mobili); in quest’ultimo caso la notevole variabilità granulometrica dei sedimenti condiziona in modo specifico l’insediamento degli organismi vegetali. Pertanto è importante conoscere le caratteristiche tessiturali del substrato mediante un’analisi sedimentologica, in modo da correlarle ai popolamenti vegetali.
I sedimenti non sono altro che il prodotto dell’azione demolitrice operata dagli agenti esogeni sulle rocce. Il vento, la pioggia, le variazioni di temperatura sono le principali cause di quest’azione che trasformano le morfologia dei territori; tramite fiumi e torrenti i sedimenti vengono trasportati verso il mare che rappresenta la loro meta finale. Il moto ondoso e le correnti di fondo influenzano sia l'erosione che la tipologia dei sedimenti. Inoltre sono presenti sedimenti di tipo organogeno formati da gusci e scheletri di organismi marini che vengono demoliti principalmente dall’azione erosiva del mare. L’analisi sedimentologica ha lo scopo di evidenziare le caratteristiche e la distribuzione dei sedimenti sui fondali e di ottenere, attraverso un’analisi statistica dei dati, informazioni circa la possibilità di movimento dei sedimenti sul fondo. La prima fase dell’analisi sedimentologica è la campionatura che deve essere effettuata in modo da prelevare un campione di sedimento indisturbato. A questo scopo viene utilizzata una benna “Shipek” formata da due semicilindri concentrici che si chiudono automaticamente quando il sistema impatta sul fondo. Essa permette in media il recupero di sedimento su un’area di circa 25x35 cm e uno spessore di circa 10 cm. Il sedimento così recuperato viene lavato in acqua dolce, per evitare la formazione di aggregati dovuta ai sali presenti, e quindi essiccato in forno. Successivamente si eseguono le analisi di vagliatura a secco con una pila di setacci a maglie via via decrescenti tali da dividere le porzioni ghiaiosa, sabbiosa e pelitica; quest’ultima viene vagliata con il metodo densimetrico. Le differenti porzioni granulometriche vengono classificate secondo la tabella di Wenthworth che divide i sedimenti nei gruppi principali (Ghiaie, sabbie, peliti) in funzione dei diametri. In seguito le percentuali delle singole porzioni vengono collocate all’interno del triangolo di Folk e Ward che presenta ai tre vertici le percentuali di ogni porzione. In tale modo è possibile definire le caratteristiche del sedimento.
Sulla base del rapporto tra specie e substrato le specie vengono raggruppate in categorie ecologiche: Epilitiche, Endolitiche, Psammofile, Pelofile, Epibionti e Endobionti.
- Le specie epilitiche vivono fissate sulla roccia che rappresenta escusivamente il supporto sul quale ancorarsi senza avere con esso nessun rapporto trofico; inoltre, è stato notato che alcune specie prediligono selettivamente substrati vulcanici, metamorfici o carbonatici. Sembra che questa preferenza sia in rapporto soprattutto con lo stato della superficie rocciosa (liscia, rugosa, finemente fessurata ecc..). Un esempio tipico Mediterraneo è quello della Rissoella verruculosa presente prevalentemente su substrati vulcanici o metamorfici.
- Le specie endolitiche sono principalmente Cyanophyceae, Chlorophyceae che vivono all’interno di roccie carbonatiche o conglomerati organogeni.
- Le psammofile sono specie che si impiantano su sabbia mediante un sistema stolonifero di ancoraggio; si tratta principalmente di Angiosperme (Posidonia, Halophila, Cymodocea) e alghe verdi (Dasycladus, Penicillus e Caulerpa).
- Le specie pelofile (Zostera e Ruppia) vivono su fango sviluppando sistemi di ancoraggio simili quelli delle specie psammofile.
- Le specie epibionti ed endobionti vivono rispettivamente su organismi o all’interno di essi utilizzandoli esclusivamente come supporto o traendo da essi sostanze organiche (parassiti o semiparassiti).
La prateria a Posidonia oceanica.
La prateria a Posidonia oceanica caratterizza il piano infralitorale su substrato mobile. Si tratta di un popolamento estremamente complesso che è stato oggetto di numerosi studi. Alcune praterie sis viluppano anche su roccia. Da un punto di vista vegetazionale, all'interno del Posidonieto si distinguono diversi aggruppamenti. Moliner (1958) distingue la biocenosi relativa alle foglie della Posidonia (Posidonietum oceanicae) e la biocenosi sciafila relativa ai rizomi (Udoteo-Peyssonnelietum). Successivamente Boudouresque (1971) riprende gli aggruppamenti descritti da Molinier (1958) anche se egli denomina Posidonietum oceanicae esclusivamente l'aggruppamento di alghe epifite della fronda. Uno studio vegetazionale specifico sulla flora epifita si deve a Van der Ben (1971).
Posidonietum oceanicae Molinier 1958
La specie caratteristica dell'associazione è: Posidonia oceanica.
In questo aggruppamento, descritto come biocenosi, Molinier (1958) individua due elementi fondamentali: la Posidonia oceanica e la componente faunistica che costituiscono, secondo l'Autore, le specie caratteristiche della biocenosi. L'Autore segnala inoltre la presenza sulle foglie di Posidonia di numerose "Melobesie incrostanti" non determinate. Nonostante le numerose specie animali presenti, la descrizione dell'aggruppamento fornita dall'Autore è estremamente esauriente e rispecchia fedelmente la fisionomia del popolamento. Pertanto riteniamo di trasformare questa biocenosi in associazione vegetale non prendendo in considerazione la componente animale e mantenendo come unica specie caratteristica Posidonia oceanica. La presenza di un'unica specie caratteristica è un criterio omogeneo a quello adottato da Pignatti (1953) per gli altri aggruppamenti ad Angiosperme marine. Quest'associazione si afferma sui substrati dell'infralitorale caratterizzati da sabbie grossolane e da ottima ossidazione; si sviluppa in maniera ottimale tra 10 e 20 metri di profondità.
Myrionemo-Giraudietum sphacelarioidis Van der Ben 1971
Le specie caratteristiche dell'associazione sono: Myrionema orbiculare, Giraudia sphacelarioides, Cladosiphon cylindricus, C. irregularis, Myractula gracilis, Chondria mairei, Spermothamnion flabellatum f. bisporum.
L'insieme delle specie epifite della fronda di Posidonia è stato considerato da diversi Autori come un aggruppamento distinto. Van der Ben (1971) ha effettuato uno studio specifico sulle specie epifite delle foglie di Posidonia oceanica delle coste francesi, identificando l'aggruppamento a Myrionema orbiculare e Giraudia sphacelarioides; la descrizione di Van der Ben accompagnata da una tabella fitosociologica esauriente rende questo aggruppamento valido come associazione vegetale che viene denominata Myrionemo-Giraudietum sphacelarioidis Van der Ben 1971. L'Autore indica come specie caratteristiche: Myrionema orbiculare, Giraudia sphacelarioides, Cladosiphon cylindricus irregularis, Myractula gracilis, Chondria mairei e Spermothamnion flabellatum f. bisporum. La presenza di queste specie sulle fronde di Posidonia è stata segnalata da altri autori; ciò testimonia la costanza di questo aggruppamento vegetale. Tuttavia questo aggruppamento non può essere considerato esclusivo delle foglie di Posidonia; in maniera impoverita infatti è presente sia sulle foglie di altre Angiosperme marine che sulla fronda di Cistoseire dell'Infralitorale.
Cymodoceetum nodosae Giaccone e Pignatti 1967
La specie caratteristica dell'associazione è: Cymodocea nodosa.
E' stata descritta dagli Autori per il Golfo di Trieste, dove la specie generalmente vicaria totalmente le praterie a Posidonia oceanica. Questa associazione, che nel Golfo di Trieste ha il suo limite inferiore a 6-7 metri di profondità, nel resto del Mediterraneo si spinge anche a 30-35 metri nell'infralitorale inferiore. Si afferma su sedimenti di sabbie con prevalenza di elementi fini scarsamente ossidati (sabbie fini ben calibrate e sabbie fangose in ambiente calmo). Gli Autori, sebbene non riportino in tabella in modo esplicito specie caratteristiche, indicano nel testo delle alghe macroscopiche osservate in questo popolamento (Gracilaria confervoides auct., Polysiphonia subtilissima).
Zosteretum marinae Pignatti 1953
La specie caratteristica dell'associazione è: Zostera marina.
Quest'associazione descritta da Pignatti (1953), occupa i fondi marini con sabbia fine mista a fango poco ossidati dell'Infralitorale superiore, in biotopi soggetti ad apporti di acque dolci e quindi con acque fresche e dissalate (22-37 %º). Si tratta di una associazione monospecifica di cui Pignatti riporta una tabella sintetica di 10 rilievi con due indici (il primo indica la copertura % ed il secondo la classe di presenza). Trattandosi di una tabella di presenza non è possibile indicare l'olotipo. Pertanto indichiamo il neotipo nel ril. n. 1 della tab. 2 in Simonetti (1973) con unica specie caratteristica Zostera marina. In questo rilievo, così come negli altri relativi all'associazione (dal 1 al 3), sono presenti anche numerose specie algali. Si tratta, nella maggior parte dei casi, di specie ubiquiste o comunque ampiamente distribuite in altre Associazioni sia algali che di altre Angiosperme marine.
Zosteretum noltii Pignatti 1953
La specie caratteristica dell'associazione è: Zostera noltii (come Z. nana)
Questa associazione predilige le velme e le paludi delle lagune caratterizzate da sedimenti fini in cui sono presenti processi riduttivi. La specie caratteristica è Zostera noltii, specie spiccatamente eurialina a ripartizione mesolitorale ed infralitorale superiore con fondali fangosi.
Si tratta di una associazione monospecifica di cui Pignatti (1953) riporta una tabella sintetica di 10 rilievi con due indici (il primo indica la copertura % ed il secondo la classe di presenza).
Trattandosi di una tabella di presenza non è possibile indicare l'olotipo; pertanto indichiamo il neotipo nel ril. 4 della tab. 2 in Simonetti 1973 con unica specie caratteristica Zostera noltii. In questo rilievo, così come negli altri relativi all'associazione (ril. 4 e 5), sono presenti anche numerose specie algali. Si tratta, nella maggior parte dei casi, di specie ubiquiste o comunque ampiamente distribuite in altre associazioni sia algali che di altre Angiosperme marine.
Syntaxa di Ordine superiore: substrato mobile
La vegetazione ad Angiosperme marine è stata oggetto di studio e di inquadramento fitosociologico da parte di numerosi autori. Braun-Blanquet (1931) inserisce nell'Ordine Potametalia Koch 1926 due Alleanze: Posidonion Braun-Blanquet 1931 e Ruppion maritimae Braun-Blanquet 1931. Nella prima Alleanza inquadra l'associazione a Posidonia e a Cymodocea, mentre nella seconda inserisce l'associazione a Ruppia e a Zannichellia pedicellata e la subassociazione a Zostera noltii.
Béguinot (1941) introdusse l'ordine Zosteretalia per la vegetazione marina ad Angiosperme.Braun-Blanquet e Tüxen (1943) modificano lo schema di Braun-Blanquet (1931) introducendo l'alleanza Zosterion Blaun-Blanquet & Tüxen 1943 al posto dell'alleanza Posidonion; essi inquadrano questa nuova alleanza nell'ordine Zosteretalia Béguinot 1941; questo viene inquadrato nella classe Potametea Braun-Blanquet & Tüxen 1943 che comprende sia le associazioni di pleustofite, che quelle delle aptofite e delle rizofite in acque salmastre e salate.
Pignatti (1953) inquadra la vegetazione marina ad Angiosperme nella classe Zosteretea Pignatti 1953. Essa, secondo l'Autore comprende la "vegetazione fanerogamica sommersa ed affiorante a bassa marea, delle acque salse". In essa Pignatti inquadra l'ordine Zosteretalia Béguinot 1941 e l'alleanza Zosterion Blaun-Blanquet e Tüxen 1943.
Tutte le Associazioni ad Angiosperme marine (Posidonietum oceanicae, Zosteretum marinae, Zosteretum noltii, Cymodoceetum nodosae) vengono pertanto inserite in questa alleanza. L'associazione Myrionemo-Giraudietum sphacelarioidis si inquadra, invece, nell'alleanza Cystoseirion crinitae. Anche l'aggruppamento vegetale ad Halophila stipulacea è compreso in questa alleanza.
La vegetazione marina dell'infralitorale su substrato mobile è inquadrata fitosociologicamente secondo il seguente schema:
- Classe Zosteretea Pignatti 1953
Car. Classe: Zostera noltii.
Tipo nomenclaturale: Zosteretalia Beguinot 1941.- Ord. Zosteretalia Béguinot 1941
Sinonimo: Zosteretalia Braun-Blanquet & Tüxen 1943.
Tipo nomenclaturale: Zosterion Braun-Blanquet e Tüxen 1943.
Car. Ord.: Zostera noltii.- All. Zosterion Braun-Blanquet & Tüxen 1943
Car. All.: Zostera noltii.
Tipo nomenclaturale: Zosteretum noltii Pignatti 1953.- Ass. Posidonietum oceanicae
- Ass. Zosteretum marinae
- Ass. Zosteretum noltii
- Ass. Cymodoceetum nodosae
- All. Zosterion Braun-Blanquet & Tüxen 1943
- Ord. Zosteretalia Béguinot 1941
La vegetazione lagunare
Per questo tipo di vegetazione gli schemi tuttora validi sono quelli elaborati da Den Hartog e Segal, i quali basano la loro classificazione su quanto precedentemente elaborato da Du Rietz, Luther e Lohmeyer. Le forme di crescita costituiscono, infatti, la base per la classificazione secondo Du Rietz; mentre Luther basa i suoi schemi sulle modalità di impianto al substrato e Lohmeyer, invece, da più importanza ai criteri di classificazione basati sul significato ecologico delle diverse specie che caratterizzano questa tipologia vegetazionale.
Il sistema di classificazione elaborato da Den Hartog e Segal è anche influenzato dalle idee di Iversen il quale nella sua classificazione fitosociologica tiene conto degli adattamenti morfologici che le piante acquatiche hanno adottato per meglio inserirsi nell'ambiente lagunare.
Secondo Giaccone e Perrera il criterio floristico, posto alla base del sistema fitosociologico di classificazione della vegetazione, va integrato con elementi di bionomia bentonica, che includono nella sistematica vegetazionale anche fattori biotici e abiotici. Inoltre, per le unità sintassonomiche superiori al rango di associazione i criteri di Du Rietz, Luther e Lohmeyer sono più capaci di esprimere le complesse interazioni dei molteplici fattori ecologici che entrano in gioco negli ambienti lagunari. Le forme di crescita e i modi di fissazione al substrato dei vegetali acquatici hanno qualche analogia, relativamente a fisionomia e struttura della vegetazione, con le forme biologiche di Raunkiaer basate sugli adattamenti dei vegetali ai periodi sfavorevoli e ai rapporti gemma/suolo.
Per quanto concerne l'inquadramento sistematico degli ambienti lagunari, Giaccone e Perrera raggruppano i vari popolamenti come segue:
a) Vegetazione di aptofite fissate sui substrati duri naturali o artificiali.
È un tipo di vegetazione del Mesolitorale e della Frangia infralitorale in cui predominano alcune specie di Enteromorpha, Blidingia e, in Alto Adriatico dove le acque sono più pulite, Fucus virsoides. Nei biotopi con buon ricambio idrico, nei mesi primaverili, lungo la frangia Mesolitorale e Infralitorale si posso sviluppare anche Petalonia fascia, Scytosiphon lomentariae e Porphyra leucosticta. Nello stesso periodo si può anche sviluppare la specie Ulva laetevirens che poi in Novembre - Dicembre è vicariata da Ulva rotundata e in Luglio - Agosto da Ulva curvata. L'Infralitorale, che data la torbidità di questi ambienti non va oltre 1 - 2 metri di profondità, è colonizzato oltre che dalle specie già citate anche da molte specie preferenziali dello Pterocladiello-Ulvetum laetevirentis. Tra queste quelle che predominano sono Chondria coerulescens, Chylocladia verticillata e Grateloupia doryphora. Nei moli e lungo le banchine di Venezia Pignatti descrive per l'Infralitorale il Cystoseiretum barbatae, un'associazione caratterizzata da un popolamento eterogeneo appartenente alla serie evolutiva che porta dal Cystoseiretum crinitae, degli ambienti puliti, allo Pterocladiello-Ulvetum laetevirentis degli ambienti con acque torbide e inquinate. In questi ambienti sono anche state riscontrate coperture notevoli di Corallina officinalis e Chondria coerulescens. Nello Stagnone di Marsala la specie Cystoseira crinita è vicariata da C. barbata f. aurantia associata ad Anadyomene stellata, Dasycladus vermicularis, Acetabularia acetabulum, Padina pavonica e C. schiffnerii. Una particolarità di questo ambiente è la presenza di C. spinosa f. marsalensis che si sviluppa con la parte stolonifera immersa nel sedimento e la parte aerea allungata fino a raggiungere la superficie dell'acqua. Confrontando i diversi biotopi lagunari si evince un progressivo impoverimento della vegetazione fino ad arrivare a popolamenti monospecifici che vanno interpretati come un complesso di facies impoverite di due associazioni presenti in condizioni ottimali. Pertanto le cinture mesolitorali presentano una facies impoverita dell'Enteromorphetum compressae, presente anche in ambienti puliti, e quelle infralitorali dello Pterocladiello-Ulvetum laetevirens, presente in ambienti instabili e/o eutrofici.
b) Vegetazione di rizofite fissate sui fondali mobili con sabbia e fango.
Prima di descrivere questi tipi vegetazionali bisogna distinguere due tipi di ambienti, quelli più tipicamente marini e quelli più tipicamente lagunari. Negli ambienti più tipicamente marini in prossimità di lagune si afferma il Cymodoceetum nodosae. Ciò accade principalmente nel Golfo di Trieste; mentre in altri biotopi mediterranei la specie Caulerpa prolifera spesso sostituisce Cymodocea e Zostera. Nello Stagnone di Marsala è possibile osservare un Posidonietum oceanicae in vari stadi di regressione. In ambienti tipicamente lagunari Posidonia e Cymodocea sono vicariate da Zostera marina, in presenza di effluenti di acqua dolce, e da Zostera-Zosterella noltii, in presenza di condizioni instabili e di substrati caratterizzati da fanghi ridotti. Negli ambienti più tipicamente salmastri e poco profondi, come quelli dalla Laguna Veneta, della Laguna di Grado, Marano e Orbetello, si sviluppano popolamenti a Ruppia spiralis. Il Ruppietum spiralis predomina anche nelle Valli di Comacchio, negli stagni di Vendicari e nelle saline di Marsala su fondali caratterizzati da limo e a profondità compresa tra i 20 ed i 150 centimetri. Negli stessi ambienti dove il substrato è prevalentemente sabbioso il Ruppietum spiralis è vicariato dal Lamprothamnietum papulosi.
c) Vegetazione di pleustofite secondarie con acropleustofite galleggianti sulla superficie dell'acqua e con bentopleustofite adagiate o rotolanti sui fondali mobili.
Le acropleustofite sono prevalentemente rappresentate da alghe verdi Ulvales con predominanza di Enteromorpha intestinalis e Ulva laetevirens. A queste possono associarsi alcune Cladophorales, tra cui predominano Cladophora, Chaetomorpha e Chondria dasyphylla. Nei periodi freddi predominano le fasi aptofitiche mentre nei periodi caldi, soprattutto in ambienti più tipicamente marini o dove le acque sono più calme, predominano le forme pleustofitiche. Nella fase acropleustifitica queste specie sembrano perdere la capacità di riprodursi sessualmente acquisendo una enorme capacità di riproduzione vegetativa, tanto da dar luogo a grandi accumuli di materia organica che a fine estate, a seguito del normale concludersi del ciclo di vita di questi vegetali, danno luogo ad intensi e distruttivi fenomeni di inquinamento organico di origine secondaria. Tutti questi popolamenti ad acropleustofite secondarie si possono considerare come lo stadio finale di una serie di trasformazioni che subisce la vegetazione marina su substrato duro in quegli ambienti, prima puliti e ben ossigenati, che subiscono trasformazioni tali che ne riducono la quantità di ossigeno disciolto e l'equilibrio ecologico in generale. Dal punto di vista ecologico hanno un notevole interesse i popolamenti caratterizzati da bentopleustofite e da mesopleustofite. Nelle lagune la specie più interessante è Gracilariopsis longissima, mentre nelle valli hanno un notevole interesse le specie Valonia aegagropila, Chaetomorpha linum e Rytiphloea tinctoria. Nello Stagnone di Marsala sono molto interessanti le ricche formazioni di Maërl con grossi sferoidi di Phymatolithon calcareum, Mesophyllum lichenoides e Lithothamnion fruticulosum auct. oltre ad altre specie di Melobesie libere.
La vegetazione di pleustofite secondarie con alghe marine acropleustofite e bentopleustofite comprende le seguenti associazioni:
Chaetomorpho-Valonietum aegagropilae Giaccone 1974
Le specie caratteristiche dell'associazione sono: Valonia aegagropila, Chaetomorpha linum.
Le specie formano egagropili sferoidali che si aggregano formando masse in aree soggette a correnti di fondo negli stagni e lagune dei vari settori del Mediteraneo.
Gracilariopsetum longissimae Giaccone 1974
La specie caratteristica dell'associazione è: Gracilariopsis longissima.
I talli di Gracilariopsis longissima formano grovigli di filamenti che, in alcuni punti si accumulano con uno spessore di oltre 50 cm. Questa associazione si sviluppa in lagune in comunicazione con il mare e soggette ad apporto di acque dolci, ricche di nutrienti.
Cladophoro-Rytiphloetum tinctoriae Giaccone 1994
Le specie caratteristiche dell'associazione sono: Cladophora echinus, Rytiphloea tinctoria.
Si tratta di un'associazione che forma popolamenti puri che si trovano su fondali sabbiosi; è stata tuttavia riscontrata come sottostrato anche nelle associazioni a rizofite e aptofite.
Queste associazioni marine non sono state dai loro autori inquadrate in unità fitosociologiche di ordine superiore. Si sviluppano nell'infralitorale superiore alto degli ambienti lagunari in comunicazione con il mare; pertanto si propone qui di inquadrare queste associazioni durevoli degli ambienti lagunari nella Classe Cystoseiretea Giaccone 1965, nell'Ordine Cystoseiretalia Molinier 1958 emend. Giaccone 1994 e nell'Alleanza Cystoseirion crinitae Molinier 1958. Nelle tabelle fitosociologiche citate, infatti, si ritrovano alcune specie eurivalenti inquadrabili negli syntaxa superiori relativi all'Infralitorale superiore. A queste specie spesso si associano come acropleustofite in primavera consorzi galleggianti di Cladophorales, di Ceramiales ed in autunno di Nostocales. Forme bentopleustofite appartengono anche alle Melobesie libere, mentre tra le alghe rizofitiche possono avere coperture significative Dasycladus vermicularis e Caulerpa prolifera.
Per gli ambienti lagunari, in letteratura, sono state descritte altre due Associazioni caratterizzate da Lamprothamnium papillosum e Ruppia spiralis. La prima specie è dominante nel Lamprothamnietum papulosi Corillion 1957 e si afferma su fondali sabbiosi in ambienti lagunari condizionati da apporto di acque dolci. L'associazione Ruppietum spiralis Iversen 1936 si afferma ad una profondità compresa tra 20 e 150 cm su fondali con predominanza di limo in ambienti salmastri (Giaccone, 1974).
L'associazione Lamprothamnietum papulosi è inquadrata nell'Ordine Charetalia Sauer 1937 e nell'Alleanza Halo-Charion Krausch 1964, mentre l'associazione Ruppietum spiralis nell'ordine Ruppietalia (Tüxen 1960) Den Hartog & Segal 1964 e nell'alleanza Ruppion maritimae (Braun-Blanquet 1931) Den Hartog & Segal 1964.
La vegetazione tionitrofila
La vegetazione fotofila di substrato duro nell'infralitorale superiore in condizioni di alterazione ambientale ed in particolare di inquinamento organico, viene vicariata da associazioni durevoli con dominanza di specie opportuniste, caratterizzate da strategia fuggitiva, ed in particolare di Ulvales. Questi aggruppamenti sostituiscono le associazioni climaciche dell'Alleanza Cystoseiron crinitae Molinier 1958 e sono, quindi, da considerarsi dal punto di vista dinamico come stadi di regressione della serie evolutiva dell'Ordine Cystoseiretalia Molinier 1958. Le specie del genere Cystoseira, infatti, costituiscono un fattore biotico fondamentale per la composizione e la strutturazione delle associazioni climaciche. Le specie opportuniste delle associazioni durevoli, infatti, regrediscono qualora vengano riequilibrati in un biotopo i fattori edafici e la serie evolutiva dei Cistoseireti riprende il suo ciclo fino a stabilizzarsi con la maturazione dell'associazione climax.
Ulvetum laetevirentis Berner 1931
Le specie caratteristiche dell'associazione sono: Enteromorpha linza, Pterosiphonia parasitica, Ulva laetevirens.
Questa associazione si sviluppa in condizioni di acqua molto calma, inquinata e diluita in maniera discontinua, con temperatura stagionalmente elevata e luminosità intensa. Si afferma su substrato compatto nella frangia infralitorale o in ambienti di pozza ed in insenature protette. Si tratta di una associazione povera in specie e ricca di individui.
L'Autore inserisce Ulva lactuca tra la specie caratteristiche di questo aggruppamento; tuttavia, la presenza in Mediterraneo di questa specie deve essere confermata. Pertanto, come proposto da Boudouresque et al. (1977) Ulva lactuca auct. è da riferire a Ulva laetevirens, che consideriamo come caratteristica anche di Ordine fitosociologico superiore.
Pterocladiello-Ulvetum laetevirentis Molinier 1958
Le specie caratteristiche dell'associazione sono: Pterocladiella capillacea (come P. pinnata), Colpomenia sinuosa, Chondracantus acicularis, Nitophyllum punctatum, Ulva laetevirens.
Le condizioni in cui si afferma questa associazione sono simili a quelle descritte per l'associazione precedente, in biotopi con una minore intensità luminosa. Tuttavia l'aggruppamento descritto da Molinier è maggiormente strutturato e caratterizzato da: Pterocladella capillacea (come P. pinnata), Colpomenia sinuosa, Chondracantus acicularis, Nitophyllum punctatum e Ulva lactuca auct. Quest'ultima specie, così come quella di Berner, è probabilmente da riferire a Ulva laetevirens.
Geraci (1990) ha studiato questo aggruppamento effettuando rilevi mensili che hanno messo in evidenza anche il periodismo stagionale e l'estrema eterogeneità dell'aggruppamento.
L'associazione definita da Molinier (1958) mostra un corteo floristico differente da quello dell'Ulvetum laetevirentis Berner. Un paragone tra le tabelle di queste due Associazioni mostra quindi che si tratta di aggruppamenti differenti in cui l'Ulva laetevirens rappresenta l'unico elemento comune in presenza di alterazione ambientale. Pertanto riteniamo Ulva laetevirens caratteristica di syntaxa di ordine superiore.
Dictyopteretum polypodioidis Berner 1931
La specie caratteristica dell'associazione è: Dictyopteris polypodioides (come Haliseris polypoidioides).
L'associazione si afferma nei primi metri dell'infralitorale in stazioni con luminosità e idrodinamismo ridotti; queste condizioni non permettono l'affermarsi di aggruppamenti vegetali più complessi e Dictyopteris polypodioides assume un ruolo predominante.
Ceramietum rubri Berner 1931
La specie caratteristica dell'associazione è: Ceramium rubrum.
Si afferma a poca profondità in biotopi con idrodinamismo e luminosità ridotti con apporto di acqua dolce. Il pieno sviluppo di questa associazione si ha in primavera.
Corallinetum officinalis Berner 1931
La specie caratteristica dell'associazione è: Corallina officinalis.
Si afferma in biotopi con idrodinamismo elevato, in condizioni di luminosità elevata e con apporto di acqua dolce.
Syntaxa di ordine superiore: vegetazione tionitrofila
Il primo a inquadrare le associazioni tionitrofile da un punto di vista fitosociologico fu Berner (1931). Questo ricercatore definisce un'Alleanza Ulvion nella quale inserisce quattro aggruppamenti: Ulvacetum, Haliseretum, Corallinetum e Ceramietum. Più tardi Molinier (1958) creò l'Ordine Ulvetalia nel quale inquadrò l'Ulvion Berner 1931 indicandone le specie caratteristiche. In questa Alleanza inserì lo Pterocladiello-Ulvetum laetevirentis Molinier 1958 considerando tale associazione un sinonimo pro parte dell'Ulvacetum Berner 1931.
Ulvetalia Molinier 1958
Tipo nomenclaturale: Ulvion laetevirentis Berner 1931.
Car. Ord.: Ulva laetevirens, Spermothamnion irregulare, Gymnogongrus griffithsiae.
Ulvion laetevirentis Berner 1931
Den. orig.: Ulvion Berner 1931.
Tipo nomenclaturale: Ulvetum laetevirentis Berner 1931.
Car. All.: Ulva laetevirens, Spermothamnion irregulare, Gymnogongrus griffithsiae.
Dall'esame comparato delle tabelle riportate dagli Autori che hanno studiato questa vegetazione riteniamo che U. laetevirens debba essere considerata anche come specie caratteristica di Alleanza e di Ordine.
La vegetazione a Caulerpe
Le alghe del genere Caulerpa sono Chlorophyceae, alghe verdi dell'ordine Bryopsidales. Di quest'ordine fa parte la famiglia Caulerpaceae costituita da un unico genere comprendente circa 72 specie raggruppate in 12 sezioni sulla base della differente ultrastruttura dei cloroplasti. Le Caulerpe sono alghe «termofile» diffuse prevalentemente nei mari caldi, tropicali e subtropicali, nei quali costituiscono a volte la parte preponderante della vegetazione algale. Il genere Caulerpa, per il momento, è rappresentato nel bacino del Mediterraneo da solo 6 specie. L'organizzazione del tallo delle Caulerpe è simile per tutte le specie in cui è suddiviso il genere. Sono, infatti, provviste di un cauloide principale strisciante ad accrescimento apicale, detto stolone, dal quale ventralmente si dipartono i rizoidi, numerosi e molto ramificati, che hanno la funzione di fissare la pianta al substrato, mentre dalla parte dorsale si alzano i filloidi, che differiscono nelle diverse specie per forma e dimensioni. Il tallo di queste alghe è costituito da cenociti privi di pareti di divisione trasversali per cui assume una struttura di tipo polienergidico. Pertanto i nuclei che formano ogni singola fronda di Caulerpa possono spostarsi in tutto il citoplasma mediante correnti plasmatiche. Data la mancanza di pareti trasversali, all'interno del tallo sono presenti numerose strutture trasversali, trabecole, che servono da sostegno all'alga per impedire che si afflosci su se stessa. Le Caulerpe si riproducono sia vegetativamente, mediante talea, che sessualmente. Nel caso della riproduzione sessuale i gameti, maschili e femminili, vengono formati per olocarpia nel medesimo tallo. Le papille riproduttive vengono differenziate nei filloidi, ma possono anche formarsi negli stipiti dei filloidi e sullo stolone.
La dispersione in Mediterraneo
Caulerpa prolifera è il più antico rappresentante di questo genere in Mediterraneo ed ha una distribuzione pantropicale. Oggi in Mediterraneo si trovano altre cinque specie di Caulerpa (C. mexicana, C. racemosa, C. scalpelliformis, C. sertularioides, C. taxifolia), tutte provenienti da mari tropicali, ed introdottesi in esso probabilmente in poco più di cento anni, cioè da quando fu tagliato il canale di Suez. Per alcune di queste specie (es. C. racemosa e C. mexicana) si pensa ad un relitto di epoca terziaria. Era stata descritta per il Mediterraneo anche un'altra specie di Caulerpa, C. olliveri, ma sembra che si tratti di un'ecoforma di prolifera ritrovata anche nei Caraibi.
Il Mare Nostrum sta infatti subendo una continua invasione da parte di organismi provenienti dal bacino Indo-Pacifico a causa di due eventi che si sono succeduti nell'arco di un secolo. Infatti, da quando, nel 1869, fu aperto Suez si è assistito ad un continuo flusso di organismi migranti, animali e vegetali, detti specie Lessepsiane, che dal Mar Rosso giungevano fino al settore Orientale del bacino Orientale del Mediterraneo. Questi organismi erano, però, confinati lungo le coste siro-palestinesi, turche ed egiziane per via del continuo afflusso di milioni di metri cubi di acqua dolce che quotidianamente il Nilo scaricava in Mediterraneo, andando a costituire una insormontabile barriera ecologica alla loro diffusione nei settori Centrale ed Occidentale del bacino Orientale. Quando nel 1965 fu chiusa la Diga di Assuan la portata d'acqua del basso corso del Nilo venne drasticamente ridotta ad un terzo diminuendo di conseguenza l'afflusso di acqua dolce in Mediterraneo. Questo diminuito apporto di acqua dolce finì così per non costituire più una barriera di bassa salinità tra i settori Occidentale ed Orientale del bacino Orientale. Inoltre le opere di escavo eseguite nel Canale dopo il 1967 hanno portato la velocità della corrente proveniente dal Mar Rosso fino a 3 - 4 nodi. Da questa data si ha, infatti, in tutto lo Jonio una invasione di specie migranti molte delle quali stazionavano da decenni sulle coste dell'Egitto, della Palestina, del Libano, della Siria, della Turchia, di Cipro ed in genere del Mare Egeo centro-meridionale. Esempi significativi di questa accelerazione espansionistica sono costituiti tra le alghe rosse da Lophocladia lallemandii, tra le alghe verdi da Caulerpa racemosa e Caulerpa scalpelliformis, tra le alghe brune da Stypopodium schimperi e tra le Angiosperme da Halophila stipulacea. Essendo molte delle Caulerpe opportuniste ed alcune possedendo anche un metabolismo eterotrofo facoltativo, mediato da batteri simbionti, le tappe dell'invasione del bacino Mediterraneo sono costituite preferenzialmente dai biotopi costieri degradati da inquinamento organico o dai margini dei Posidonieti, caratterizzati da abbondanza di resti vegetali e di materiale organico proveniente dall'ecosistema prateria. Ciò è dimostrato dal fatto che il ricoprimento delle praterie a Caulerpa diminuisce, quando la sostanza organica viene ossidata o metabolizzata dagli organismi in situ oppure quando se ne ha un abbattimento mediante gli impianti di depurazione. Le specie migranti con portamento invasivo (C. mexicana, C. racemosa, C. scalpelliformis, C. sertularioides, C. taxifolia), più o meno favorite da azioni antropiche, hanno trovato in Mediterraneo un'area biogeografica ospitale anche a causa dei processi di tropicalizzazione delle sue acque e di meridionalizzazione dei suoi settori settentrionali. Lüning (1990) a ragione inseriva il Mediterraneo nella fascia calda dell'oceano mondiale.
Distribuzione in Mediterraneo
Caulerpa prolifera è presente pressoché lungo tutte le coste del bacino come è ampiamente riportato in letteratura e raggiunge la profondità di oltre cento metri in Mediterraneo Orientale e di circa quaranta metri in quello Occidentale. Le Caulerpe di recente introduzione nel Mediterraneo, provenienti dal Mar Rosso, sono presenti prevalentemente nel settore Orientale e Centro-Orientale del bacino. Infatti, fu proprio l'estremo settore Orientale di questo mare il loro primo punto di arrivo. Da quì seguendo la direttrice di migrazione Est-Ovest si sono poi diffuse a tutto lo Jonio e l'Adriatico e stanno ora spingendosi verso il Mediterraneo Occidentale. Non si può escludere che alcune popolazioni siano derivate da esemplari provenienti dall'Atlantico tropicale e subtropicale (es. C. racemosa delle coste tunisine). Nel Mediterraneo Orientale erano segnalate, infatti, già nella prima metà del secolo oltre a Caulerpa prolifera anche C. mexicana, C. scalpelliformis e C. racemosa. Più recentemente è stata segnalata C. sertularioides. C. racemosa ha avuto il maggior successo tra tutte. Infatti, dopo aver raggiunto le coste della Tunisia, ad Ovest, e dell'Egeo, ad Est, è riuscita a giungere sino alle coste di Lampedusa e Augusta (Sicilia Orientale). Attualmente è segnalata in Adriatico, lungo le coste albanesi, e nel Mediterraneo Occidentale, in Toscana e in Sardegna (località Sarroch-Cagliari; Marco Giani, com.pers.). C. mexicana è presente lungo le coste di Siria e Palestina. C. scalpelliformis e C. sertularioides per il momento sono segnalate solo nel Bacino Orientale del Mediterraneo, la prima lungo le coste di Libano, Siria, Palestina e nel Mar Egeo, e la seconda nel Mar di Marmara, in Turchia. C. scalpelliformis nel 1998 è segnalata in Sardegna meridionale. taxifolia fu segnalata nel 1984 nelle acque del Principato di Monaco. Da allora taxifolia si diffuse molto rapidamente andando a colonizzare ampi tratti di mare lungo la Costa Azzurra ricoprendo migliaia di ettari di fondale. Nel 1992 fu segnalata la presenza di C. taxifolia anche alle Baleari, in Spagna, e lungo i litorali ligure e toscano, in Italia. Recentemente (inverno 1995) è stata segnalata anche in Adriatico sulle coste della Croazia davanti Spalato. Attualmente si sta diffondendo in Sicilia orientale e meridonale. A proposito delle tre specie C. mexicana, C. scalpelliformis e C. taxifolia in letteratura si è ipotizzato che il complesso mexicana-taxifolia sia un unico taxon in fase di speciazione attiva. Riteniamo che nel complesso si possa includere anche C. scalpelliformis. Pertanto riteniamo fondata l'opinione che queste tre entità siano ancora in fase di speciazione, soprattutto in condizioni di simpatria.Effetti della dispersione delle Caulerpe sulla vegetazione e l'ambiente naturale del Mediterraneo
L'arrivo in Mediterraneo di specie tropicali a portamento invasivo del genere Caulerpa ha causato, in qualche caso, effetti rilevanti sulla vegetazione marina (alghe e fanerogame) delle località interessate. C. prolifera trovandosi in Mediterraneo da sempre si è ben integrata nella vegetazione di questo mare divenendo una componente normale del fitobenthos. La sua vegetazione è inclusa nella serie dinamica della prateria a Posidonia. Per la specie sertularioides, ancora confinata all'estremo settore Orientale di questo bacino con una distribuzione pressoché puntiforme (Mar di Marmara), in base alla scarsa letteratura esistente a tutt'oggi non può dirsi nulla circa gli effetti che la sua presenza può avere avuto sulla flora algale del luogo in cui è stata segnalata. Questa segnalazione necessita di ulteriore conferma.
C. scalpelliformis, che ha una distribuzione più estesa (lungo tutto il litorale siro-palestinese), è stata oggetto di studi fitosociologici da parte di Mayoub (1976), il quale ha riscontrato la presenza di circa 20 specie epibionti per ciascun rilievo effettuato, con un valore medio di R/P pari a 5,11. Inoltre, dalle tabelle che l'A. riporta non si evincono effetti dannosi sui preesistenti popolamenti. Nel Golfo di Cagliari la specie è attualmente confinata davanti all'area industriale di Sarroch.
C. mexicana, presente in Mediterraneo Orientale e si trova mescolata a tutta la vegetazione fotofila dell'infralitorale superiore.
Gli studi condotti sul popolamento a racemosa di Baia S.Panagia (Sicilia Orientale) hanno messo in evidenza come un fondale pressoché privo di vita possa essere ricondotto sulla via del risanamento ambientale grazie all'insediamento di una specie vegetale in grado di colonizzare ampi tratti di mare fortemente degradati dagli insediamenti industriali. La racemosa di Baia S.Panagia si è, infatti, insediata in un tratto di mare nel quale il massivo apporto di inquinanti (sversamento di idrocarburi, scarichi di acque di centrali termiche, scarti di lavorazioni industriali in genere) avevano portato a morte, prima della realizzazione degli impianti di depurazione, un preesistente Posidonieto, del quale oggi restano soltanto i rizomi ricoperti da uno spesso strato di sabbia grossolana e fango, con il conseguente azzeramento della biodiversità. I valori degli indici sinecologici riscontrati per questo popolamento sono: 46,75 specie vegetali epibionti in media per rilievo mentre il valore medio dell'indice R/P è di 8,12.
Un discorso più articolato va fatto per C. taxifolia. Questa Cloroficea, infatti, dal momento della sua comparsa in Mediterraneo ha destato le preoccupazioni del mondo scientifico internazionale per via della sua elevata velocità di colonizzazione su qualunque tipo di substrato. In Costa Azzurra la sua espansione fece temere per la vita stessa dei preesistenti Posidonieti, già spesso in precarie condizioni per il degrado ambientale e per le condizioni idrologiche sfavorevoli, al punto da far si che si attivassero diversi gruppi di ricerca nel tentativo di mettere a punto metodologie mirate all'eliminazione dell'alga.
Boudouresque et al. (1994) affermarono che Caulerpa taxifolia costituisce una minaccia per la diversità biologica dell'intero Mediterraneo. Verlaque & Boudouresque (1995) successivamente dichiararono che la flora epifita di taxifolia è fortemente danneggiata e ridotta. Da ricerche da noi condotte sulla biodiversità degli epibionti vegetali nei prati a Caulerpa taxifolia nel Principato di Monaco sono venuti risultati molto diversi da quanto asseriscono gli A.A. citati. Gli indici sinecologici mostrano valori senzaltro più confortanti rispetto a quelli calcolati per la vegetazione epifita associata a due fanerogame (Cymodocea nodosa e Halophila stipulacea) viventi nel Mediterraneo. Per la flora epibionte, infatti, abbiamo rilevato un numero medio di specie per campione pari a 30 unità ed un valore medio dell'indice R/P pari a 8,64. Questi dati, che in valore assoluto sono inferiori a quelli calcolati per gli epifiti di Posidonia oceanica, sono però, senzaltro, paragonabili, in quanto a diversità biologica, con quelli calcolati per la fanerogama tenendo conto della minore superficie di impianto offerta agli epibionti dai filloidi di C. taxifolia. Solo in estate-autunno si nota una certa diminuzione del numero di specie per rilievo dovuta all'intensa attività metabolica della Clorofita. Da osservazioni recenti si è potuto constatare come l'espansione di taxifolia sia in fase di rallentamento in alcune zone interessate da fioriture di alghe mucillaginogene. Ciò probabilmente sta a significare che la specie può aver raggiunto una certa integrazione con la flora preesistente, e con gli organismi animali che se ne cibano. In pratica esisterebbero specie vegetali ed animali in grado di competerle gli spazi vitali.
Furono presi in esame anche i metaboliti secondari prodotti da C. taxifolia come possibili agenti in grado di sopprimere oltre che gli altri vegetali marini anche gli animali che avessero pascolato l'alga. Qualcuno ipotizzò che anche l'uomo, nutrendosi di tali organismi (pesci e molluschi) avrebbe potuto averne conseguenze dannose. Infatti, nel genere Caulerpa, tra i metaboliti secondari si trovano principalmente caulerpenina, un sesquiterpenoide acetilenico con un legame triplo e quattro legami doppi; caulerpina, un pigmento derivato da un dimero dell'indolo e caulerpicina, una miscela di composti simili agli sfingolipidi dei tessuti animali. I prodotti considerati più tossici sono quelli di natura terpenoidica, gli altri hanno funzione soprattutto antibiotica, detterrente-gustativa e allelopatica o sociologica. Composti simili sono però presenti anche in altre alghe (Flabellia petiolata, Halimeda tuna, Anadyomene stellata, Cystoseira sp. pl., ecc.) ampiamente diffuse in Mediterraneo. Da ciò si evince che tali deterrenti chimici sono sempre stati attivi nei rapporti tra pascolanti e pascolo e risulta dunque spropositato affermare che possano essere responsabili di eventi patologici per le specie animali, uomo compreso. Secondo Vidal et al. (1984), inoltre, caulerpina e caulerpicina hanno scarsa o nulla tossicità. Molte specie di Caulerpa sono coltivate e mangiate nell'area del Pacifico.
In conclusione, si può affermare che sui substrati mobili senza vegetazione i prati a Caulerpe, sia indigene che esotiche, non costituiscono un danno per la biodiversità algale, ma anzi una opportunità di ripopolamento di aree degradate. Per cui la dispersione in Mediterraneo di specie Indo-Pacifiche o in genere ad affinità calda costituisce un fenomeno di ricolonizzazione di un bacino geologicamente e biologicamente giovane caratterizzato dall'esistenza di biotopi con spazi disponibili e di biocenosi sociologicamente aperte. Gli inconvenienti segnalati o temuti sia per le praterie ad Angiosperme marine che per la vegetazione algale di substrato duro sono dovuti al carattere invasivo che molte specie dimostrano nell'attuale fase di dispersione in Mediterraneo. Ma fenomeni simili sono segnalati nell'Eocene e nell'Oligocene dell'Europa centrale per via indiretta dalla presenza diffusa di Molluschi erbivori obbligati delle Caulerpe. L'accentuazione dei fenomeni migratori nel bacino orientale, fanno di questo relitto di mare Tetide un ponte biogeografico verso l'Indo-Pacifico, che nell'ultimo secolo è stato riattivato con il taglio di Suez ed è stato potenziato con l'incremento del traffico mercantile in questa via d'acqua artificiale. Ci sembra che si possa ragionevolmente affermare che l'attuale fase di dispersione invasiva della vegetazione a Caulerpe in Mediterraneo, sia l'effetto di cause complesse ad azione sinergica come ad esempio:
- eccessivo degrado della fascia costiera,
- intensificazione del traffico marittimo,
- opere di escavo del Canale di Suez dopo il 1966,
- chiusura della Diga di Assuan dopo il 1965,
- diffuse pratiche di acquariologia e di maricoltura,
- aumento dei valori medi nella temperatura dell'acqua di mare (dati del «World Climate Disc»; 1992),
- probabili fenomeni evolutivi con meccanismi epigenetici di origine ambientale nei popolamenti a Caulerpe presenti in Mediterraneo.
Le Caulerpe, secondo la teoria di Hutchinson (1959), conosciuta come «Omaggio a Santa Rosalia», negli ambienti in equilibrio si comportano a nostro avviso come "compattatrice di nicchia"; negli ambienti stressati esse assumono, invece, un ruolo di risanamento e di incremento della biodiversità. L'attuale successo nella fitness moltiplicativa delle specie del genere Caulerpa in Mediterraneo può essere spiegato non con lo sciovinismo nazionalista o l'isterismo ambientalista, ma soltanto con i concetti acquisiti dalla moderna ecologia: «É una buona ipotesi di lavoro predire che il climax non arriverà mai e che qualunque comunità è sotto lo stress della colonizzazione». Pertanto non si comprende perché debbano fare eccezione le comunità a Posidonia e a Cystoseira.
Ci sembra di potere concludere questo argomento con la spiegazione biogeografica che Lüning (1990) dà del successo della colonizzazione delle specie migranti o introdotte in Mediterraneo: «A causa della relativa recente ricolonizzazione del biota marino intorno a circa 5 milioni di anni, il Mediterraneo può manifestare una più grande ricettività alle specie migranti o variamente adattate di quanto ci si potrebbe aspettare per un antico, evoluto ecosistema nel quale lo spazio è scarso e la competizione alta». Le comunità biologiche sono definite dagli habitat; gli habitat del Mediterraneo negli ultimi 60 anni sono cambiati sia per cause meteomarine che biotiche anche di origini antropiche, di conseguenza cambiano le comunità. Alla fine del secondo millennio è il momento delle comunità a Caulerpe, ma non è la fine del mondo, anzi del Mediterraneo, ma un'altra tappa nella storia multiforme della vita in questo travagliato habitat del Pianeta Terra.
Le Associazioni vegetali a Caulerpe
Caulerpetum racemosae Giaccone & Di Martino 1995
La vegetazione a racemosa è di tipo psammofilo. Le rizofite dominanti sono, oltre alla specie caratteristica, anche Caulerpa prolifera alla quale si associano in sottostrato Rytiphloea tinctoria e Halopteris filicina. Più a Nord (Brucoli) e più a Sud (Ognina di Siracusa) C. racemosa si ritrova mescolata sia alla vegetazione epilita che a Posidonia oceanica.
I prati si presentano con una struttura orizontale a mosaico o in densi prati continui. La struttura verticale è costituita da un'associazione dipendente in epibiosi e da specie emisciafile in sottostrato, che consideriamo riferibili alla classe Lithophylletea Giaccone 1965. Nella sezione delle specie epibionti ci sembra opportuno segnalare la presenza delle seguenti specie che solo occasionalmente si ritrovano in altri tipi di vegetazione: Microdictyon tenuius, Champia parvula, Halodyction mirabile e Lejolisia mediterranea. Le quattro specie possono essere indicate come caratteristiche elettive dell'associazione dipendente in epibiosi sull'associazione principale rappresentata dalla sezione delle rizofite.
La vegetazione di sottostrato è relativamente povera di specie e senza significativa capacità di ricoprimento. Si tratta di specie incrostanti o meno a larga ripartizione in sottostrato o infeudate agli aggruppamenti sciafili del sistema fitale.
La vegetazione a racemosa possiede carattere invasivo ed è in rapida espansione in Mediterraneo. La sua fascia climatica di diffusione è tutta l'area tropicale.
Ci sembra che lo studio fitosociologico effettuato porti alla individuazione di una nuova associazione rizofitica (1) e di una nuova associazione dipendente epifitica (2).
1) Caulerpetum racemosae Giaccone & Di Martino 1995
rilievo tipo: ril. n°2 tab. 2 sezione specie rizofite
profondità: -5 m,
luogo classico: Baia di S.Panagia - Siracusa - Sicilia, esposizione E, pendenza (°) = 10
specie caratteristica: Caulerpa racemosa (Forsskäl) J.Agardh
condizioni ambientali: infralitorale sabbioso e alquanto infangato in aree moderatamente riparate.
2) Microdictyetum tenuii Giaccone & Di Martino 1995
rilievo tipo: ril. n°2 tab. 2 sezione specie epifite su racemosa
profondità: -5 m
luogo classico: Baia di S.Panagia - Siracusa - Sicilia, esposizione E, pendenza (°) = 10
specie caratteristiche elettive: Microdictyon tenuius (C.Agardh) Decaisne, Champia parvula (C.Agardh) Harvey, Halodictyon mirabile Zanardini e Lejolisia mediterranea Bornet
condizioni ambientali: associazione dipendente di epibionti su Caulerpa racemosa in aree moderatamente riparate nell'infralitorale.
La validità della prima associazione è confortata dalla consolidata prassi di riconoscere il rango di associazione alla vegetazione marina rizofitica con contingente caratterizzante monospecifico. La validità delle associazioni dipendenti ed in particolare di quelle epifitiche nella vegetazione marina, è stata riconosciuta per l'aggruppamento vivente sulle foglie di Posidonia oceanica. In conclusione si può affermare che la fedeltà degli epibionti in questa tipologia vegetazionale deve essere considerata solo elettiva e non esclusiva. Ma studi ulteriori su altri tipi di vegetazione epifitica marina ci sembra debbano essere intrapresi prima di tentare, ove possibile, un inquadramento sintassonomico di queste associazioni dipendenti in unità di ordine superiore.
Caulerpetum taxifoliae mexicanae Giaccone & Di Martino 1995
Nel biotopo studiato la vegetazione a C. taxifolia ecad mexicana è fissata su un ghiaione grossolano originato dalle correnti dello Stretto di Messina. Il suo modo di fissarsi al substrato la rende idonea ad essere classificata sia come rizofitica che come aptofitica. La propagazione avviene mediante stoloni unicamente basali. La struttura orizzontale è a chiazze mentre quella verticale è nettamente divisa in più sezioni. Nella sezione rizofitica è nettamente dominante C. taxifolia ecad mexicana, mentre Halopteris filicina ha solo un'elevata presenza. Sulla specie dominante si sviluppa in epibiosi un'associazione dipendente altamente diversificata, ma anche con evidenti valori di ricoprimento dei filloidi e degli stoloni. Consideriamo caratteristiche elettive tra gli epifiti Aglaothamnion byssoides, Crouania attenuata e Laurencia microcladia. I maggiori valori di ricoprimento sono presentati da Dasya rigidula, Dictyota dichotoma v. intricata, Hydrolithon farinosum, Lithophyllum pustulatum e Laurencia minuta ssp. scamaccae.
Nella sezione delle aptofite viventi sia in sottostrato che più raramente in strato elevato presentano alta frequenza e significativo ricoprimento Dictyota dichotoma v. dichotoma, Nithophyllum punctatum, Lithophyllum frondosum, Peyssonnelia rosa-marina e stagionalmente il gametofito di Asparagopsis armata. In questa sezione si affollano un numero elevato di specie rare o occasionali. Probabilmente l'elevata reologia della colonna d'acqua e l'instabilità del substrato mobile favoriscono le efemeroficee sulle alghe perennanti. Ma il ruolo stabilizzante di mexicana potrebbe in futuro cambiare questo rapporto sopratutto nell'orizzonte basso dell'infralitorale superiore.
A conclusione dello studio fitosociologico effettuato sono state descritte due nuove associazioni di alghe marine una per la sezione rizofitica (1) e l'altra per quella epifitica (2).
1) Caulerpetum taxifoliae mexicanae Giaccone & Di Martino 1995
rilievo tipo: ril. n°6 tab.1 sezione specie rizofite
luogo classico: Stretto di Messina, litorale Torre Faro-Ganzirri, profondità -4m su substrato di ghiaia e ciottoli; esposizione S-SE, pendenza (°) = 10.
specie caratteristica: Caulerpa taxifolia (Vahl) Agardh ecad mexicana Sonder ex Kützing sensu Coppejans et al. (1990 e 1992).
condizioni ambientali: ambiente leggermente eutrofico moderatamente riparato dal moto ondoso, con reologia intensa, unidirezionale oscillante. Infralitorale superiore.
Riteniamo di distinguere la nostra associazione dal Caulerpetum scalpelliformis Mayhoub 1976, perchè le due specie presentano ecologia e distribuzione batimetrica differente. Inoltre C. scalpelliformis non è ancora arrivata in Sicilia, anche se è già (1998) segnalata in Sardegna.
2) Laurencietum microcladiae Giaccone & Di Martino 1995
rilievo tipo: ril.n°6 tab.1, sezione specie epifite su C. taxifolia ecad mexicana
profondità -4 m;
luogo classico: Stretto di Messina, litorale Torre Faro-Ganzirri, esposizione S-SE, pendenza (°) = 10.
specie caratteristiche: Laurencia microcladia Kützing
elettive: Aglaothamnion byssoides (Arnott ex Harvey in W.J.Hooker) L'Hardy Halos et Ruenes e Crouania attenuata (C.Agardh) J.Agardh.
condizioni ambientali: reologia intensa, eutrofia moderata. Infralitorale superiore.
Le ragioni che giustificano la descrizione delle nuove associazioni vegetali sono simili a quelle riportate in un lavoro contestuale sulla vegetazione a Caulerpa racemosa e quindi non intendiamo qui riportarle. In ogni modo il nostro procedimento metodologico ripercorre quello canonico del trattato di fitosociologia di Braun-Blanquet (1964), adottato in precedenza per la vegetazione marina dell'infralitorale.
Caulerpetum scalpelliformis Mayoub 1970 emend. Giaccone & Di Martino 1995
Come corollario, infine, intendiamo validare ed emendare l'associazione cumatofila, termofila e fotofila di substrato mobile descritta come popolamento a Caulerpa scalpelliformis da Mayhoub (1976). In accordo, infatti, con il codice di nomenclatura fitosociologica (Barkman et al., 1986), è necessario indicare il rilievo tipo, non riportato nella descrizione originale dall'autore. Le specie caratteristiche dell'associazione sono: Caulerpa scalpelliformis (Brown ex Turn.) C. Agardh, Schizotrix arenaria (Berkeley) Goumont. Nell'emendamento consideriamo specie caratteristiche Caulerpa scalpelliformis e Schizotrix arenaria (riportata con il sinonimo Microcoleus chtonoplases). Caulerpa mexicana, al dire dello stesso autore, si associa a C. scalpelliformis solo negli ambienti più riparati e superficiali dell'infralitorale superiore, mentre quest'ultima specie forma estesi e densi prati fino a 40 m di profondità.
Caulerpetum scalpelliformis Mayhoub 1976 emend.Giaccone & Di Martino 1995
Den.orig.: Peuplement à Caulerpa scalpelliformis
lectotipo: rilievo stazione n°70 (Tartous) tab. n°20
tab.20: 4 rilievi dell'A.; loc.: Siria; data 1973; sup. ril. cm2 900; pend. (°): da 0 a 45; esp.: varia; prof. cm: da 60 a 100;
specie caratteristiche: Caulerpa scalpelliformis (Brown ex Turn.) Agardh, Schizotrix arenaria (Berkeley) Gomont.
Caulerpetum taxifoliae taxifoliae Di Martino & Giaccone 1996
La vegetazione a taxifolia può impiantarsi su vari tipi di substrato, ma ha una maggiore capacità di colonizzazione su substrato mobile o su roccia in ambienti soggetti ad intensa sedimentazione.
La vegetazione dominante sui substrati di sabbia grossolana sotto la gettata del Museo, prima dell'insediamento di C.taxifolia, era la prateria a Posidonia oceanica. Le mattes morte con rizomi ancora evidenti, sono visibili in alcune aree di erosione. Il Posidonieto però, è morto prima dell'arrivo della Caulerpa a causa della instabilità ambientale, causata in tutta la regione dai lavori di riempimento del mare sia del Principato di Monaco che davanti all'aeroporto di Nizza. Notevole influenza hanno avuto gli scarichi fognari non depurati e le acque meteoriche non dissabbiate.
I prati a C.taxifolia si presentano con una struttura orizzontale omogenea, interrotta quà e là da chiazze con fronde e stoloni putrescenti soprattutto alla fine dell'estate. Su alcuni spuntoni di roccia emergente dal prato dominano Dictiotali e Sfacelariali, che rappresentano la vegetazione vestigiale di substrato roccioso.
La struttura verticale è costituita da una associazione rizofitica, con una sola specie dominante ( taxifolia) e due specie accompagnatrici (Halopteris filicina e Stypocaulon scopariae). Sulla rizofita dominante si sviluppa una flora epifilla ed in parte epifita sul cauloide, dotata di una considerevole biodiversità. Una sessantina di specie sono state trovate solo come epibionti ed una quarantina comuni anche alla vegetazione del sottostrato.
Esclusivamente nel sottostrato abbiamo trovato solo 16 specie, tutte dotate di scarsissima capacità di ricoprimento, analogamente alla maggior parte delle specie epibionti. Questa struttura è paragonabile alla vegetazione rizofitica con dominanza di Caulerpe sia autoctone (prolifera) che alloctone (scalpelliformis, mexicana, racemosa, ecc.).
Ci sembra che lo studio fitosociologico effettuato porti alla individuazione di una nuova associazione rizofita, già rilevata, ma non formalmente descritta secondo il Codice di Nomenclatura Fitosociologica (C.N.F.), da Verlaque & Fritayre (1994).
Si tratta di una vegetazione la cui validità fitosociologica è confortata dalla consolidata prassi, accettata dal C. N.F., di riconoscere il rango di associazione alla vegetazione marina rizofitica con contingente caratterizzante monospecifico.
Contrariamente a quanto affermato da Verlaque et al. (1994, 1995) e da Boudouresque & Tripaldi (1994): "L'epiflore de taxifolia est negligeable", a noi la flora epifita risulta consistente e ci sembra di poter riconoscere una nuova associazione dipendente in epibiosi caratterizzata, come la specie supporto, da due specie algali anch'esse recentemente migrate in Mediterraneo: Acrothamnion preissii e Womersleyella setacea.
La validità delle associazioni dipendenti ed in particolare di quelle epifitiche nella vegetazione marina è stata riconosciuta per l'aggruppamento vivente sulle foglie di Posidonia oceanica ed è comunemente riconosciuta nella vegetazione terrestre di tipo arboricolo.
rilievo tipo: n° 3 tab. 2 sezione specie rizofite;
profondità: -16 mt.; pendenza: 10%;
luogo classico: gettata del Museo Oceanografico Principato di Monaco;
specie caratteristica: Caulerpa taxifolia (Vahl) C. Agardh ecad taxifolia sensu Coppejans et al. (1990 e 1992).
condizioni ambientali: infralitorale fotofilo sabbioso-ciottoloso con peliti in ambiente moderatamente calmo.
rilievo tipo: n° 3 tab. 2 sezione specie epifite;
profondità: -16 mt.; pendenza: 10%;
luogo classico: gettata del Museo Oceanografico Principato di Monaco;
specie caratteristiche elettive: Acrothamnion preissii (Sonder) Wollaston Womersleyella setacea (Hollenberg) R.E.Norris
condizioni ambientali: associazione dipendente di epibionti su Caulerpa taxifolia, ma rinvenibile impoverita anche su altri substrati vegetali e minerali, infralitorali, caratterizzati da instabilità di sedimentazione e da ambienti moderatamente calmi.
Caulerpetum proliferae Giaccone & Di Martino 1997
Ollivier (1929: 103) descrive la velocità di diffusione della vegetazione a C.prolifera sulla Costa Azzurra con espressioni forti e preoccupate che ricordano quelle più recenti degli algologi e dei giornalisti francesi per C.taxifolia: «Il devient tellement envahissant qu'en général là où il s'installe toute autre végétation disparait sur le fond». Ma nel 1977 Meinesz sperimentava colture in situ con Caulerpa prolifera per ripopolare i fondali della regione Alpes-Maritimes.
Caulerpetum proliferae Giaccone & Di Martino 1997
Olotipo: ril. 4, Tab. 2
Tab. 2: 8 ril. in Giaccone & Di Martino 1997;
Luogo classico: Capo S. Croce (Sicilia orientale, Siracusa);
Data: 1996; sup. ril. cm2: 3.600; pend: 10 %; esp.: S-SE; prof.: 5 e 10 mt.
Specie caratteristica: Caulerpa prolifera (Forsskäl) Lamouroux
Condizioni ambientali: infralitorale fotofilo, sabbia grossolana con peliti in ambiente moderatamente calmo.
La vegetazione epifitica comprende prevalentemente le componenti lessepsiane riscontrate nel Caulerpetum taxifoliae (Acrothamnietum preissii) ma vi sono presenze significative anche delle associazioni dipendenti ritrovate negli altri Caulerpeti della Sicilia orientale. Questo risultato era in qualche modo atteso, perchè Caulerpa prolifera è il taxon più anticamente insediato e più diffuso in Mediterraneo. Inoltre la vegetazione rizofitica associata sia in strato elevato che in sottostrato ha una biodiversità maggiore di quella osservata nei Caulerpeti delle specie migrate o introdotte dagli ambienti tropicali degli oceani vicini.
Syntaxa di ordine superiore
Classe Caulerpetea Giaccone & Di Martino 1997.
Classe di rizofite algali associate ai prati formati da specie del genere Caulerpa. L'ambiente di elezione è l'Infralitorale, ma può estendersi anche al Circalitorale degli ambienti tropicali e subtropicali. Pur potendosi sviluppare su differenti substrati, le Caulerpe prediligono i substrati clastici con importante componente di sostanza organica, che utilizzano con un metabolismo fotochemio-organotrofo e con un efficiente meccanismo di assorbimento rizoidale. La vegetazione è generalmente eurifotica, euriterma e galenofila.
- Classe Caulerpetea Giaccone & Di Martino 1997.
Tipo nomenclaturale: CAULERPETALIA Giaccone & Di Martino 1997.
Car. class.: Caulerpa prolifera e Caulerpa racemosa- Ordine: CAULERPETALIA Giaccone & Di Martino 1997.
Tipo nomenclaturale: CAULERPION Giaccone & Di Martino 1997.
Car. ord.: Caulerpa prolifera e Caulerpa racemosa- Alleanza: CAULERPION Giaccone & Di Martino 1997.
Tipo nomenclaturale: Caulerpetum racemosae Giaccone & Di Martino 1995
Car. all.: Caulerpa prolifera e Caulerpa racemosa
- Alleanza: CAULERPION Giaccone & Di Martino 1997.
- Ordine: CAULERPETALIA Giaccone & Di Martino 1997.