1. Elementi di corologia della biodiversità vegetale marina del Mediterraneo
La corologia è la scienza che studia la distribuzione attuale delle specie. Sulla base delle affinità filogenetiche indaga, inoltre, sull'origine, sulla ecologia dell'evoluzione e sugli eventi che hanno originato i fenomeni della vicarianza e della dispersione dei singoli taxa o di interi sintaxa.
La corolgia è in grado di evidenziare, con metodi biologici, fenomeni geodinamici, come la tettonica a placche, e fenomeni paleoclimatici e le conseguenti crisi di salinità, di temperatura, di trofia ecc..., dei bacini marini ed in particolare del Terziario e del Quaternario.
L'origine della biodiversità sul Pianeta Terra: modelli a confronto
Nel 1959 Hutchinson, uno dei fondatori della scuola ecologica americana, pubblicò la sua teoria sull'origine della biodiversità che aveva in precedenza esposta in un Congresso mondiale a Palermo. L'articolo dell'American Naturalist è così titolato."Homage to Santa Rosalia or why are there so many kinds of animals?" Certamente nel dedicare questa teoria, rimasta ancora un caposaldo nell'ecologia dell'origine delle specie, l'ecologo statunitense volle esprimere il suo stupore davanti alle tante teofanie che Palermo evoca, vista da Monte Pellegrino: ricchezza di natura e di cultura, simbolicamente sintetizzate nella grotta - santuario della "Santuzza" palermitana, che nell'800 aveva riempito di ammirato stupore anche Goethe.
Secondo il modello di Hutchinson l'esistenza di molteplici nicchie ecologiche (intese come un ipervolume di assi di esigenze, di risorse e di habitat) spiega perché vi sono molteplici forme di vita, cioè tante specie.
La competizione interspecifica per le risorse nelle comunità naturali ha selezionato nel tempo evolutivo le specie e ne ha reso possibile la coesistenza attraverso una gamma infinita di mutualismi, di divergenza di caratteri e di comportamenti. Secondo questa teoria la diversità nelle comunità è una ricchezza da condividere e chi non condivide le risorse e non modera la competizione si estingue.
Un modello semplice di questa teoria è stato elaborato nel 1972 da Mac Arthur.
Una seconda teoria sulla consistenza della biodiversità nelle comunità è stata formulata nel 1978 da Connel, stimolato da studi sulle foreste pluviali e sulle barriere coralline.
Egli formulò l'ipotesi che le catastrofi naturali (uragani e tifoni per le foreste, fenomeni meteo - marini come "el Niño" per le barriere, incendi spontanei per praterie e boschi), liberando le nicchie dai vecchi occupanti, ringiovaniscono gli ecosistemi mantenendone alta la diversità biologica.
Questa teoria è stata denominata del "disturbo intermedio" e si basa ormai su una grande raccolta di dati sperimentali, che dimostrano l'esistenza dei maggiori picchi di diversità non nei sistemi in equilibrio o climacici, non nei sistemi degradati da inquinamento, ma nelle comunità sottoposte a moderata instabilità a causa sia di fattori biotici (erbivoria, predazione) sia di fattori abiotici (instabilità climatica, idrodinamica, edafica).
Una terza teoria generale sulla biodiversità fu formulata tra il 1966 e nel 1981 da Paine e riorganizzata nel 1993 da Colinvaux, che la denominò "Principio del raccolto". Si basa su numerose sperimentazioni in natura, che evidenziano una maggiore diversità nelle comunità dove vi sono più "raccoglitori di risorse": vi sono più specie vegetali, dove vi sono più erbivori che impediscono che pochi vegetali forti competitori occupino tutto l'habitat; vi sono più erbivori nei sistemi naturali dove vivono più specie di carnivori. Viceversa la diversità diminuisce drasticamente se crollano i carnivori terminali ed intermedi.
Tutte queste teorie, integrandosi danno una risposta ragionevole sul perché della diversità nella biosfera.
I fattori che regolano la biodiversità in Mediterraneo.
Il Mediterraneo rappresenta soltanto lo 0,8% della superficie marina dell'Oceano mondiale; ma la consistenza della sua biodiversità è paradossalmente relativamente elevata. Si calcola che in tutti gli oceani ed i mari del mondo gli Animali marini (Metazoi) sono circa 130.000 taxa ed i Vegetali (Macrofitobenthos) circa 8.000 taxa.
In Mediterraneo di questi Animali sono presenti circa 7.241 taxa, pari al 5,5% (Fredj et al., 1992); mentre dei Vegetali se ne contano circa 1.351 taxa, pari al 16,2% (Giaccone et al., 1999). (Tab. 1 e 2).
Secondo il modello di Mac Arthur (1972 in Begon et al., 1989), che qui applico per il Mediterraneo, vi sono in un ecosistema più specie per i seguenti motivi:
- a) una maggiore gamma di risorse (oligotrofia diffusa);
- b) gli organismi sono più specializzati (endemismi e specie stenovalenti);
- c) ciascuna specie ha una nicchia ecologica più sovrapposta con le nicchie vicine (mutualismo e specie accompagnatrici);
- d) l'asse o l'ipervolume di risorse è più sfruttato, cioè la comunità di organismi è più satura in relazione alla portanza biologica del sistema (articolati rapporti trofici e struttura biocenotica elevata).
In effetti il Mediterraneo è un mare oligotrofico e non è omogeneo né nell'habitat né nelle biocenosi e si articola in due bacini: occidentale ed orientale; ciascun bacino si divide in settori o mari minori (Alboran; Ligure; Balearico; Tirreno: alto, medio e basso; Adriatico: alto, medio e basso; Jonio: alto, orientale, occidentale, Mare Africano; Egeo: centrale, settentrionale, Mare Levantino, ecc.).
Gli endemismi specializzati per le nicchie mediterranee sono circa un terzo della sua biodiversità: 28,6% Animali e 26,6% Vegetali.
Nello studio delle biocenosi, inoltre, le specie, che accompagnano gli elementi caratteristici, sono molto numerose, perché sono in grado di sovrapporre per una elevata percentuale i loro spazi o volumi di nicchia ecologica, abbassando la competizione interspecifica ed esaltando le sinergie mutualistiche (Bellan-Santini et al., 1994; Giaccone & Di Martino, 1997).
L'ipervolume di risorse non è ugualmente ed intensamente sfruttato nei differenti bacini e settori nei quali si articola la biogeografia ed il diacronismo evolutivo del Mediterraneo.
La biodiversità e la strutturazione biocenotica diminuiscono drasticamente procedendo dal Mediterraneo occidentale al Mare Adriatico ed al Mediterraneo orientale (Tab. 3). Per quanto concerne, inoltre, la biodiversità vegetale, in Egeo si riduce fino al 32,2 % (Athanasiadis, 1987).
Origine della diversità vegetale in Mediterraneo.
Tra i fattori che hanno prodotto l'attuale biodiversità del Mediterraneo ne illustrerò brevemente due: 1) la tettonica delle placche ed in particolare alcuni eventi geodinamici, che hanno interessato nelle diverse ere geologiche i rapporti tra i corpi idrici dell'Atlantico e dell'Indo-Pacifico; 2) le variazioni climatiche negli ultimi 30.000 anni, che hanno causato le "crisi di Sapropel".
Lo studio sulla distribuzione degli organismi presenti in Mediterraneo, ha consentito di costruire uno spettro corologico, dal quale partiamo per investigare la loro origine, cioè il probabile diacronismo del loro processo evolutivo.
Approfondendo recentemente (Giaccone et al., 1999) lo spettro corologico della flora marina bentonica delle coste italiane(Tab. 4), che rappresenta il 72,76 % di quella nota per l'intero Mediterraneo, abbiamo ottenuto alcuni risultati che ci hanno fatto formulare alcune ipotesi di lavoro sull'origine della diversità vegetale marina, estensibili all'intero Mediterraneo (Tab.5 - 6 - 7).
I 202 taxa vegetali, pari a 22,32% compresi negli elementi biogeografici comuni agli oceani Atlantico e Indo-Pacifico (AP; IA; IAt; IP; P) ed in particolare quelli presenti nella fascia tropicale e subtropicale, possono essere considerati elementi tetidei e mesogei. Alcuni (IP) sono rimasti probabilmente in parte in rifugi marini marginali anche durante la crisi evaporitica del Messiniano superiore, altri rientrarono in Mediterraneo durante alterne vicende di comunicazione, susseguitesi sia durante che dopo il Terziario superiore, anche fino al Pleistocene inferiore, tra il Mediterraneo e l'Indo-Pacifico.
Alcune specie degli elementi endemici (M: 25,64%) sono molto antichi e si devono considerare veri paleoendemsimi (5%) risalenti (Stam et al., 1988; Giaccone e Di Martino, 1997) all'Oligocene (Laminaria rodriguezii, Posidonia oceanica, Rissoella verruculosa, specie del genere Rodriguezella, ecc.) o al Miocene inferiore e medio (Cystoseira corniculata, C.barbata, Halophila stipulacea, ecc.). Altri elementi sono più recenti ed alcuni costituiscono neoendemismi (21%) risalenti ad epoche successive alla trasgressione pliocenica dei Trubi, quando si riformò l'ambiente marino del Mediterraneo attuale (altre specie di Cystoseira, specie di Laurencia, ecc.).
A questo stesso evento sono legati gli elementi plio-pleistocenici (254 taxa, pari al 27,06 %) comuni soltanto all'Atlantico ed al Mediterraneo (A; Ab; Abt; At;).
Gli elementi cosmopoliti (217 taxa, pari a 23,97 %) comuni al Mediterraneo, all'Atlantico, all'Indo-Pacifico o diffusi un po' ovunque nelle zone marine del globo (Aptf; Iatf; SC; C; CA; CB; CBA), si possono fare risalire a specie distribuite in tutto l'Oceano mondiale (o in alcune sue fasce climatiche) nel corso delle ere geologiche e disperse attraverso vie marine e durante eventi generalmente poco noti.
Le variazioni climatiche avvenute durante il Pleistocene inferiore e l'Olocene hanno prodotto 12 crisi di Sapropel negli ultimi 30.000 anni (Stanley, 1978)
Il regime pluviale nei bacini che alimentano i fiumi che sboccano nel Mar Nero, determinano con un ritorno di circa 3.000 anni, crisi anossiche di origine bosforica nell'Egeo.
Il regime monsonico dell'Africa orientale, legato ai cicli di Milankovicth, causa alluvioni nilotiche e crisi anossiche nello Ionio con conseguenze negative fino alle coste della Sicilia e nei casi più gravi porta ad una inversione delle correnti generali del Mediterraneo.
Secondo recenti osservazioni oceanografiche riportate da Doumenge (1993) una crisi di Sapropel è già in atto e dovrebbe avere il suo effetto devastante intorno al 2020 - 2050.
Già nell'Egeo, infatti, sono state trovate aree profonde con acqua anossica di origine bosforica ed in molti settori dell'Adriatico e del Tirreno la risalita in superficie delle correnti di Levante hanno alterato i rapporti nelle acque tra N ed il P (Orel et al., 1993). Come conseguenza si sono avute nell'ultimo decennio varie fioriture algali, denominate "mucillagini".
Queste crisi climatiche ricorrenti sono probabilmente responsabili (Por et al., 1985) della scarsa biodiversità animale (40%) e vegetale (32%) e della destrutturazione delle biocenosi ed in particolare delle associazioni vegetali in Egeo ed in tutto il Mediterraneo orientale, che presenta nicchie ecologiche semivuote ed esposte ad invasioni di specie opportuniste alloctone e/o fuggitive (specie del genere Caulerpa, degli Ordini Ceramiales e Dictyotales: Ribera e Boudouresque,1995; Giaccone & Di Martino, 1997).
Meccanismi di diversificazione evolutiva.
La successione batimetrica dei paesaggi marini sulle coste rocciose del Mediterraneo è evidenziata nell'ambiente sommerso da formazioni vegetali, caratterizzate nello strato elevato da specie del genere Cystoseira. Il portamento della fronda e le strutture di ancoraggio al substrato di queste specie sono il risultato di una evoluzione guidata da un parametro abiotico della nicchia ecologica: l'idrodinamismo.
Le Cistoseire simpodiali (C.amentacea, C.brachycarpa, C.barbatula, ecc.) occupano le zone caratterizzate da correnti multidirezionali e bidirezionali con effetti dirompernti, mentre le specie monopodiali (C.mediterranea, C.elegans, C.sauvageauana, C.spinosa, ecc.) caratterizzano le zone con correnti deboli unidirezionali o formano isole di vegetazione nelle pozze calme e nelle baie riparate.
In ambienti con rocce organogene o vulcaniche irregolari ed in ambienti con reologia intensa le specie hanno evoluto un elaborato sistema di ancoraggio fatto da lunghi e suddivisi ramponi, terminati da ventose digitate (C.jabukae, C.tamariscifolia, C.brachycarpa v.claudiae, C.zosteroides, C.usneoides, ecc.)
In ambienti coperti da sedimenti, le specie hanno sviluppato cauloidi striscianti e corti rami primari (C.corniculata, C.dubia, ecc.).
All'interno di questa selezione evolutiva le specie hanno contrastato l'eccesso di sfruttamento provocato dall'erbivoria, elaborando sostanze allelopatiche. Queste molecole nelle specie più antiche sono semplicemente metaboliti secondari non lipofili mentre in quelle mediamente evolute sono diterpenoidi lineari, che diventano sempre più complessi con nuclei eterociclici, man mano che le Cistoseire colonizzano nuove nicchie ecologiche, nelle quali devono limitare la predazione di nuovi o più numerosi erbivori.
È stato possibile ricostruire anche su basi chimiche (Piattelli, 1990; Amico, 1995) la filogenesi delle specie attraverso la biogenesi dei prodotti allelopatici. Un esempio di speciazione simpatrica ancora non completata è stata studiata in alcuni ibridi naturali di C.algeriensis x C.elegans, caratterizzati da metaboliti a biogenesi mista (Amico et al., 1988).
In base ai risultati di questa indagine chemiotassonomica e tenendo presente la biogeografia delle differenti specie del genere Cystoseira, ritengo di poter indicare in Cystoseira corniculata, specie confinata in Adriatico, in Egeo e nello Jonio orientale ed ancora presente nell'Alto Oceano Indiano, come il taxon che ha inventato i metaboliti secondari diterpenoidi lineari come sostanze allelopatiche antierbivore.
Questa specie risale probabilmente al Mare Mesogeo del Tortoniano e rimase rifugiata durante la crisi messiniana nei bacini marginali e residuali della Paratetide e dei laghi-mare dell'Egeo. La metilazione di gruppi laterali di molecole terpenoidiche complesse è il mezzo più elaborato, espresso dai genomi più evoluti delle specie di questo genere, che hanno colonizzato aree superficiali o sono state per lunghi periodi sottoposte ad intenso idrodinamismo in nicchie, dove è anche maggiore e più diversificata la pressione degli erbivori.
Una causa di speciazione allopatrica è l'isolamento geografico, che alcune specie vegetali mediterranee hanno subito in seguito al realizzarsi di areali disgiunti. Qui indicherò soltanto due esempi riferibili ad eventi geodinamici dell'Oligocene e due esempi che si possono fare risalire una al bacino Paratetide e l'altro alla trasgressione pliocenica dei Trubi (Fig. 1 e 2).
Laminaria rodriguezii è un paleo-endemismo che ha affinità con due specie dello stesso genere (L.sinclairii e L.longipes) oggi presenti sulle coste settentrionali del Pacifico tra il Giappone, le isole Kurili, l'Alaska e la California. La via marina ipotizzata, percorsa dalla specie ancestrale, è quella medio-asiatica (Via Turgai), che collegava nell'Oligocene un Mare Artico temperato con il Pacifico settentrionale e la porzione centrale del Mare Mesogeo. Laminaria rodriguezii mantiene ancora le esigenze ancestrali: termiche (temperature inferiori ai 15°C), fotiche (luminosità bassa) e reologiche (correnti di fondo); queste condizioni si realizzano soltanto in alcuni ambienti del Circalitorale in tutto il Mediterraneo. Anche la datazione biologica basata sulla percentuale di ibridazione genica (Stam et al., 1988) ancorché approssimata, porta all'Oligocene. Alla stessa epoca si può fare risalire l'origine di Posidonia oceanica, ma attraverso il ramo orientale e meridionale della Tetide e della Mesogea. Oggi il genere si presenta con specie disgiunte in Mediterraneo e nell'Australia.
Il Fucus virsoides dell'Adriatico è rifugiato allo sbocco di acque dolci sulle coste rocciose dall'Albania a Venezia e mostra maggiori affinità con il Fucus spiralis dell'Atlantico settentrionale (Mare del Nord), che con le popolazioni delle Canarie e delle coste europee dell'Atlantico subtropicale.
Il bacino della Paratetide, attivo soprattutto nel Terziario inferiore e medio è stato la culla evolutiva di questa specie.
Laminaria ochroleuca presente nello Stretto di Messina è l'esempio più recente di disgiunzione di areale nel Mediterraneo. Questa specie è presente fino a circa 20 metri di profondità nell'area Lusitano-Senegalese dell'Atlantico e fino al Sud Africa. In Mediterraneo continua la sua distribuzione nel Mare di Alboran e fino in Algeria. La popolazione dello Stretto di Messina, per la sua profonda penetrazione batimetrica (tra -30 e -95 m) e per la sua attività fotosintetica (Drew, 1972), è già formata da una specie fisiologica, distinta dalla popolazione atlantica, dalla quale è isolata almeno da 2-3 milioni di anni. Questo isolamento e questo inizio di differenziamento evolutivo sono condivisi con altre specie vegetali (Cystoseira usneoides, Saccorhiza polyschides) ed animali (es. Errina aspera e Pachylasma giganteum) presenti nelle biocenosi bentoniche dello Stretto di Messina (Di Geronimo, 1995; Fredj e Giaccone, 1995).
Conclusioni
Nel 1820, cioè quasi all'inizio dell'impatto grave sull'ambiente causato dalla civiltà industriale, che persegue un modello di economia individualista e massimizzante, Giacomo Leopardi, con il pessimismo che lo caratterizzava scriveva in un suo pensiero:"... tanto è possibile che l'uomo viva staccato affatto dalla natura dalla quale sempre più ci andiamo allontanando, quanto un albero tagliato dalla radice fiorisca e fruttifichi. Sogni e visioni. A riparlarci di qui a cent'anni. Non abbiamo ancora esempio nelle passate età, dei progressi di incivilimento smisurato, e di uno snaturamento senza limiti. Ma se non torneremo indietro, i nostri discendenti lasceranno questo esempio ai loro posteri, se avranno posteri" (Zibaldone, 18 - 20 Agosto, 1820; Ed. 1898-1900).
L'umanità, in effetti, ne ha riparlato a Rio de Janeiro nel 1992 e a Kjoto nel 1997, per concludere che stiamo praticando uno sviluppo socio-economico non sostenibile dalla Natura e che stiamo alterando profondamente l'aria che respiriamo ed il clima che condiziona la vita nella biosfera.
Si ritiene che la nicchia ecologica, che ha reso possibile l'emergere dell'Homo sapiens, circa centomila anni fa, sia stata l'area geografica dell'Africa meridionale compresa tra il Kenia e l'Uganda, caratterizzata ancora oggi a terra e a mare da una straordinaria e spettacolare biodiversità. L'attrazione, che genera nell'uomo il fascino della biodiversità, rivela forse un imprinting, che la nostra specie ha ricevuto nella primordiale nicchia ecologica, dalla quale è emersa nella linea evolutiva dei Primati su disegno eterno di Dio-Evolutore, come Lo chiama Teilhard de Chardin (1955) fondatore dell'antropologia moderna. Più sommessamente mi sembra di poter supporre, inoltre, che il fascino che ci attira verso i luoghi ricchi di biodiversità, i boschi, i fondali marini, ma che ha spinto anche i migliori urbanisti a creare nelle città ville, giardini, arredo a verde di vie e piazze, sia forse più profondamente la nostalgia del Giardino di Eden dove, metà dell'umanità, che si riconosce nella fede di Abramo (Ebraismo, Cristianesimo ed Islamismo), crede che i nostri progenitori abbiano appreso, passeggiando e conversando con Dio, a conoscere la biodiversità del Pianeta Terra ed il segreto del suo "divenire-creatore" cioè del processo evolutivo.
La dimensione ecologica è presente soprattutto nelle nuove generazioni. Anche la grande passione per il mare, che è esplosa tra le generazioni di fine millennio, è voglia di natura incontaminata, di spazio dove potersi isolare, immergendosi e risentire, come all'inizio, che sulle acque aleggia lo Spirito di Dio, mentre la terra va ridiventando, per il degrado antropico e più recentemente per i frequenti incendi dolosi, "una massa senza forma e deserta" (Genesi, 1 e 2).
L'aumentata sensibilità ambientalista, l'interesse per gli studi ecologici sulla biodiversità nelle nuove generazioni, hanno spinto anche i nostri Corsi di Laurea ad attivare insegnamenti e/o indirizzi di biologia ed ecologia terrestre e marina. Tutto questo è forse un riemergere, nelle coscienze di fine millennio, della voce suadente dell'Eden che ricorda all'uomo di non avere avuto in possesso ma in custodia la biodiversità del Pianeta, anche al fine di coltivarla e trasmetterla alle generazioni future.
Su basi scientifico-filosofiche questa istanza interiore, ha ricevuto negli ultimi decenni una prima formulazione di una ipotesi cosmologica dell'evoluzione in Carter (1974). Il suo "Principio antropico", infatti, postula (nell'emergere nell'uomo di una forma di vita che prende coscienza del processo evolutivo del cosmo) la causa finale, che da senso ai valori delle "costanti universali" ed alla struttura della materia, che caratterizza sia il microcosmo che il macrocosmo (Rondinara, 1991).
In questi ultimi trent'anni tutte le nazioni stanno facendo a gara per vincolare almeno il 20% del territorio a parchi e riserve, detti anche "santuari della Natura". In questo linguaggio ritorna la sacralità delle religioni naturali: nel santuario della Natura, l'umanità forse ha nostalgia della esperienza dell'Eden, quando Dio scendeva a passeggiare con l'uomo, che così ne sperimentava la familiarità tra la biodiversità del Giardino.
Un richiamo autorevole alla coscienza ecologica dell'umanità, al fine di perseguire la solidarietà intergenerazionale, viene da Giovanni Paolo II (1979 e 1987):"Non bisogna fare impunemente uso delle diverse categorie di esseri viventi o inanimati (animali, piante, elementi naturali) come si vuole a seconda delle proprie esigenze economiche..., è necessario rispettare l'integrità ed i ritmi della natura e tenerne conto nella programmazione dello sviluppo...; per non mettere seriamente in pericolo la loro disponibilità non solo per la generazione presente, ma soprattutto per quelle future... Il mare è veramente un dono: è l'erario della città. L'eredità da trasmettere integra ai figli... Il mare è tra le realtà della natura che più parlano all'animo umano, chiamandolo a guardare oltre, a elevarsi in alto".
Fonti principali:
- Giaccone G. & Di Martino V. (1997) - Syntaxonomic relationship of the mediterranean phytobenthos assemblages: paleoclimatic bases and evolutive tendencies. Lagascalia 19 (1-2): 129-144.
- Giaccone G. (1999) - Ecologia della biodiversità in Mediterraneo. Accademia Internazionale di Scienze e Tecniche Subacquee; Quaderno n° 22: 43-62.
- Giaccone G. (1999) - L'origine della biodiversità vegetale del Mediterraneo. Notiziario SIBM; 35: 35-51.
Fig 1 - Comunicazioni tra Mari e Oceani durante il Cretaceo superiore ed il Terziario inferiore.
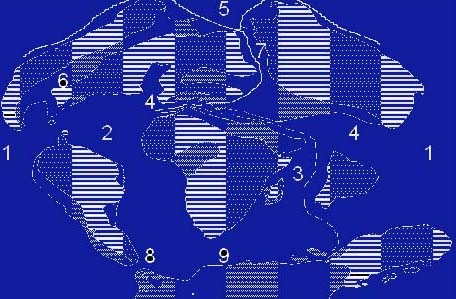
6: Via Marina Cannonbal, 7: Via Marina Turgai, 8 e 9: Vie Marine Australi (secondo Sims, 1990).
Probabili vie di dispersione delle specie ancestrali di Laminaria rodriguezii , di Posidonia oceanica, di Cystoseira corniculata, C. barbata e di Rodoliti (Corallinales)... (secondo Giaccone & Di Martino, 1997).
Fig 2 - Laghi-mare e Paratetide durante il Terziario medio e superiore.
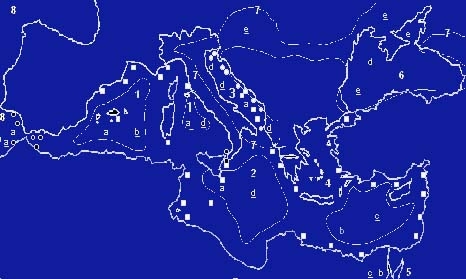
Distribuzione attuale della vegetazione relitta a Fucus virsoides (Paratetide), Cystoseira tamariscifolia, Cystoseira usneoides, Laminaria ochroleuca (Trasgressione Pliocenica), e a Caulerpa sp.pl. (Giaccone & Di Martino, 1994). Le lettere indicano i valori di salinità attuale; quelle sottolineate riportano i valori presunti nell'ambito dei laghi-mare durante il Messiniano: a = 33-38‰, b = 40-60‰, c = 70-100‰, d = 15-30‰, e = 8-12‰. (Lo schema è stato elaborato in base ai dati paleontologici e sedimentologici: secondo Por et al., 1985 e secondo Giaccone & Di Martino, 1997).
Tabella 1 - Biodiversità degli organismi marini
| Superficie | Vegetali (Macrofitobentos) | Animali (Metazoi) | |
|---|---|---|---|
| Oceano mondiale | 99,2 % | 8.000 | 130.000 |
| Mediterraneo | 0,8 % | 1.351 (16,2 %) | 7.241 (5,5 %) |
Tabella 2 - Spettro corologico degli organismi marini nel Mediterraneo
| Animali | Vegetali | |
|---|---|---|
| Mediterraneo (endemismi) | 28,6 % | 26,6 % |
| Mediterraneo + Atlantico | 50,1 % | 48,6 % |
| Mediterraneo +Indo-Pacifico | 4,4 % | 4,9 % |
| Mediterraneo + Atlantico+ Indo-Pacifico | 16,8 % | 19,9 % |
Tabella 3 - Organismi mariniriscontrati negli ultimi cinquantta anni in tre settori del mediterraneo
| MediterraneoOccidentale | MareAdriatico | MediterraneoOrientale | |
|---|---|---|---|
| Animali sp. n° 4094 /7.241 | 87,1 % | 48,9 % | 43,1 % |
| Vegetali sp. n° 1.100 /1.351 | 89,5 % | 64,4 % | 57,6 % |
Tabella 4 - Spettro corologico della flora marina bentonica delle coste italiane.
| Elementibiogeografici | N° taxa | % |
|---|---|---|
| A (Atlantico) | 13 | 1,44 |
| Ab (Atlantico boreale) | 181 | 20,00 |
| Abt (Atlanticoboreo-temperato) | 39 | 4,30 |
| At (Atlantico tropicale) | 21 | 2,32 |
| AP (Atlanto-Pacifico) | 23 | 2,54 |
| Aptf (Atlanto-Pacifico temperatofreddo) | 10 | 1,10 |
| IA (Indo-Atlantico) | 100 | 11,05 |
| IAt (Indo-Atlanticotropicale) | 6 | 0,66 |
| Iatf (Indo-Atlanticotemperato freddo) | 8 | 0,90 |
| IP (Indo-Pacifico) | 45 | 4,97 |
| P (Pantropicale) | 28 | 3,09 |
| SC (Subcosmopolita) | 96 | 10,60 |
| C (Cosmopolita) | 91 | 10,05 |
| CA (Circumaustrale) | 1 | 0,11 |
| CB (Circumboreale) | 8 | 0,90 |
| CBA (Circumboreoaustrale) | 3 | 0,33 |
| M (Mediterraneo) | 232 | 25,64 |
| Totale taxa (esclusi 78 Taxa inquirenda) | 905 |
Tabella 5 - Origine della flora marina bentonica delle coste italiane.
| Elementi | N° taxa | % |
|---|---|---|
| tetidei e mesogei | 202 | 22,3 |
| plio - pleistocenici | 254 | 28,0 |
| cosmopoliti | 217 | 23,9 |
| endemici mediterranei | 232 | 25,6 |
Tabella 6 - Composizione e spettro rispettivamente della flora marina bentonica delle coste italiane (inclusi 78 taxa inquirenda) e del Mediterraneo. (± = valori approssitivamente stimati; R/F = indice biogeografico Rhodophyceae/Fucophyceae}.
| Coste italiane | Mediterraneo | |||
|---|---|---|---|---|
| N° taxa | % | N° taxa | % | |
| Rhodophyceae | 540 | 54,94 | ± 795 | 58,84 |
| Fucophyceae | 205 | 20,86 | 265 | 19,61 |
| Chlorophyceae | 173 | 17,60 | 214 | 15,84 |
| Cyanophyceae | 56 | 5,70 | ± 56 | 3,70 |
| Xanthophyceae | 2 | 0,20 | ± 2 | 0,14 |
| Monocotyledones | 5 | 0,50 | 5 | 0,37 |
| Incertae sedis | 2 | 0,20 | ± 14 | 1,03 |
| Totali | 983 | R/F = 2,63 | ± 1351 | R/F = 3,00 |
Tabella 7 - Composizione della Flora marina bentonica delle coste italiane (inclusi 78 taxa inquirenda) compresa nelle aree di pesca FAO. (3. Tirreno e altri bacini; 4. Adriatico; 5. Ionio; 3 - 5. segnalazioni non precesate per le coste siciliane; R/F = indice biogeografico Rhodophyceae/Fucophyceae}.
| R/F | 2,79 | 3,06 | 2,98 | 3,08 |
| Aree di pesca FAO | 3 | 4 | 5 | 3 - 5 |
| Rhodophyceae | 489 | 353 | 460 | 74 |
| Fucophyceae | 175 | 115 | 154 | 24 |
| Chlorophyceae | 146 | 121 | 109 | 21 |
| Cyanophyceae | 40 | 28 | 27 | 4 |
| Xanthophyceae | 2 | - - | 1 | - - |
| Monocotyledones | 4 | 4 | 3 | - - |
| Incertae sedis | 9 | 6 | 5 | 1 |
| Taxa totali | 865 | 627 | 759 | 124 |
| % su 983 taxa | 87,99 | 63,78 | 77,21 | 12,62 |